Part of my intent for this chapter is to remove
some of the "mystery" from how the brain works. I want to convey a
sense that the brain functions like a "machine," and that living
things are automatons, consistent with this book's reductionist approach.
The human brain consists of a primitive
hindbrain, a small mid‑brain section, and a large and complicated forebrain.
The mid‑brain has a minuscule function, and won’t
be described here.
The cerebral cortex, comprising 70% of human
brain volume, consists of a left and right cerebral hemisphere, with an interconnecting
corpus callosum. Although the cerebral cortex is only 1/8‑inch thick, its
surface area is about 1 ½ square feet, and it has evolved a folded
configuration to allow the surface to fit within the human skull. The inside
surface of the cortex (gray matter) has an immense number of nerve fibers
(white matter) providing connections to other parts of the cortex, the limbic
system and other brain components.
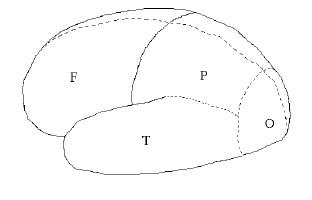
Figure 7.01. Brain lobes: Frontal, Parietal, Temporal, Occipital. View is of the left side, front is toward the left.
The corpus callosum (not shown) connects all
four lobes of one side to the corresponding lobes on the other side. This nerve
bundle is located underneath the frontal and parietal lobes, at about the same
level as the temporal lobe.
The part of the parietal lobe bordering the frontal lobe, area P1 in Fig. 7.02, is the "sensory cortex" (or "somatic cortex"). This strip of cortex is where in situ sensory signals from the body arrive. Next to the somatic cortex P1 is an area, F1, located in the frontal lobe and called the "motor cortex" or "motor strip." The motor cortex issues commands for movement (requests, actually, since sub‑cortical regions may "veto" the requests).
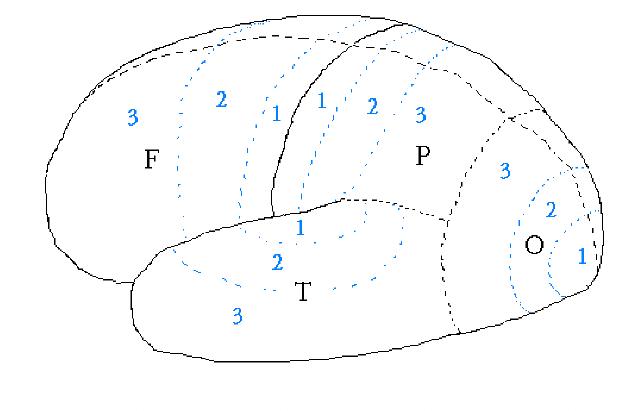
There's a one‑to‑one mapping of body location to
position along the sensory cortex strip, P1, and motor cortex strip, F1. Starting
from the part of the strips closest to the center‑line (top of brain) and going
outward, body positions are allocated in the following sequence: leg, neck, head, arm, elbow, etc, to face,
lips, teeth and tongue.

For example, when something is "felt" the flow of nerve activity "flows" according to the depiction of the figure above. When something is "heard" the flow of nerve activity "flows" according to the depiction of the following figure.
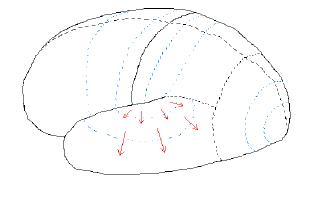
Figure 7.04. Flow of nerve activity when
something is "heard."
|
|
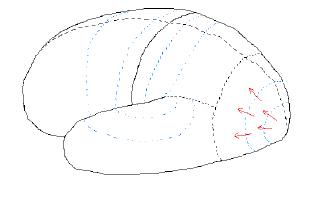
For each posterior lobe the pattern of nerve activity is the same: primary activity leads to secondary activity, which then leads to tertiary activity. The next step is for tertiary activity in adjoining areas to “compare notes,” or interact with each other.
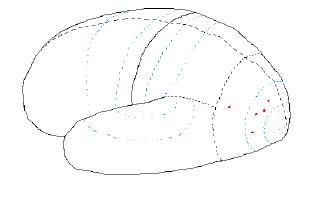
Figure 7.06. Nerve activity when a "coffee
cup" is seen.
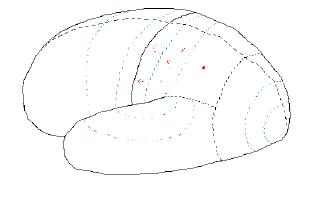
Figure 7.07. Nerve activity when a "coffee cup" is felt.
The coffee cup may be heard, as it is set down on a table. In this case activity will occur in the temporal lobe, such shown in the next figure.
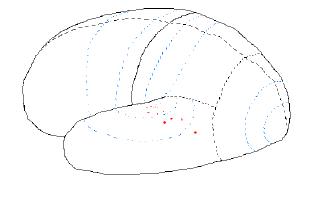
Figure 7.08. Nerve
activity when a "coffee cup" is heard, as for example being set down
upon a table.
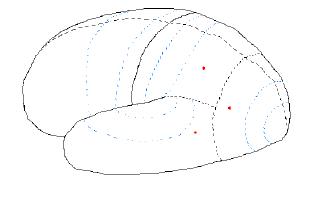
Figure 7.09. Nerve
activity corresponding to "coffee cup."
|
|
Figure 7.10. Flow of nerve activity when some
activity is planned and performed. The flow in this case is from tertiary to
primary.
The frontal lobe architecture is analogous to
that of the posterior lobes, in that the most conceptualized of ideas and plans
are created in the frontal tertiary cortex, which delivers vague "executive"
directives to the frontal lobe's secondary cortex, which formulates more
specific action commands and delivers them (as necessary) to the motor strip. The
motor strip requests permission from the sub‑cortical "reticular
activating system" (RAS), and if the RAS approves the request it is acted
upon by sub‑cortical brain areas (Luria, 1973), which carry out specific
actions (orchestrated in detail by the cerebellum).
Laterality
Proto‑humans probably had left/right symmetry,
in the sense that the right and left cerebral hemispheres had identical
capabilities, being mirror images of each other in layout. This would have provided redundancy in case one
side was injured (by a fall or blow to the head). Modern humans have asymmetric
brains: the left and right cerebral hemispheres, LB and RB, are somewhat
different, and are "specialized" for certain types of tasks. RB has
more long‑distance inter‑connections than LB, whereas LB has many areas that
are highly intra‑connected, which in turn are connected to other highly intra‑connected
regions within LB.
The best known of LB's highly intra‑connected
areas are Wernicke's Area (language comprehension) and Broca's Area (language
production). Wernicke's Area is located near the interface of the three
posterior lobes, in LB only (right-most pattern of dots in Fig. 7.11, upper).
Broca's Area is located in the frontal lobe's secondary cortex, in LB only (left-most
pattern of dots in Fig. 7.11, upper). There's a discernible pattern for the
tasks performed in these specialized, highly intra‑connected LB areas: namely, these tasks are inherently
sequential, which means that the temporal order of events is crucial! For
example, both receptive and productive language involves the processing of
sequential events (sound perception and production). Changing word order can
profoundly change meaning ("Ed ate the bear" versus "The bear
ate Ed."). In contrast, RB tasks are holistic; they resemble those that a
parallel computer processor (neural network) performs, such as instantaneous
image recognition.
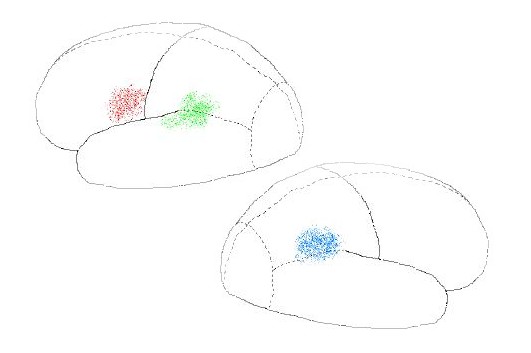
It is interesting that RB's counterpart to
Wernicke's Area, shown in Fig. 7.11 (lower panel) is devoted to the task of
monitoring the location of body parts in relation to each other and the immediate
physical environment. This area, called the "inferior parietal
lobule," or IPL, plays a critical role during manual interactions with the
environment, such as reaching out to pick fruit from a nearby branch.
It is tempting to conjecture that before humans
were capable of speech the left hemisphere's IPL counterpart region also
functioned like the present‑day IPL in RB. Because reaching out to pick fruit
had sequential components, it would have been natural for mutations to modify
what once was an LB IPL in a way that later presented an opportunity for
further modification that led to a simple form of language capability. This
region must have been built-upon to produce our present‑day Wernicke's Area,
which plays a critical role in language comprehension. This task consists of
monitoring the relationship of sound percepts to each other over time, somewhat
similar to the way the RB's IPL monitors body part relationships over time. As
Wernicke's Area evolved in LB, it must have gradually displaced the former IPL
function.
Traditional psychologists must have resented the
newcomers to their field who used instruments to measure things, and who used
rigorous techniques to study long-standing matters that had been the subject of
arm chair speculation. The old-fashioned psychologists accused those who
studied split brain patients, and found LB and RB differences, as suffering
from “dichotomania” – as if the new investigators were over-interpreting their
data due to an excess of enthusiasm. But the data is convincing, and often
dramatic.
Occasionally, a patient whose corpus callosum
has been cut can still manage to communicate in a simple way using the
rudiments of grammar. These cases offer very interesting insights into the
differing "personalities" of LB and RB. One famous example was
reported by Gazziniga (1978) which suggests that LB and RB can have different
goals in life. Their oft‑used subject P.S. was questioned about his job choice
in an experiment that allowed only RB to answer, and "automobile
race" was spelled out. As Gazziniga writes "This is most interesting,
because the left hemisphere frequently asserts that he wants to be a
draftsman" (p. 143). How poignant!
Two problems are presented simultaneously, one to the talking left brain and one to the non-talking right brain. The answers for each problem are available in full view in front of the patient. Gazzaniga and LeDoux (1978).

Figure 7.12. “Chicken claw experiment.” The “task” (top) has two parts, presented to a brain half. The answer choices, below, are in full view to both brain halves.
...the experiment requires each hemisphere to
solve a simple conceptual problem. A distinct picture is lateralized to
one hemisphere: in this case, the left sees a picture of a claw. At the
same time the right hemisphere sees a picture of a snow scene. Placed in front
of the patient are a series of cards that serve as possible answers to the
implicit questions of what goes with what. The correct answer for the left
hemisphere is a chicken. The answer for the right hemisphere is a shovel.
After the two
pictures are flashed to each half-brain, the subjects are required to point to
the answers. A typical response is that of P.S., who pointed to the chicken
with his right hand [controlled by
the left brain] and the shovel with the left [controlled by the right
brain]. After his response we asked him, "Paul, why did you do
that?" Paul looked up, and without a moment's hesitation said from
his left hemisphere, "Oh, that's easy. The chicken claw goes with the
chicken and you need a shovel to clean out the chicken coop."
It is hard to
describe the spell-binding power of seeing such things.
My interpretation is that the normal brain is
organized into modular-processing systems, hundreds of them or maybe thousands,
and that these modules can usually express themselves only through real action,
not through verbal communication. Gazzaniga (1985).
Frontal
Lobes
This was dramatically illustrated by the famous
case of Phineas Gage, who suffered a railway construction accident in 1848 that
caused a metal tamping rod to explosively penetrate and destroy his LBF (and a
small part of RBF). Without the inhibiting effect of LBF upon RBF, his behavior
was "fitful, irreverent, indulging at times in the grossest profanity...
at times pertinaciously obstinate... he has the animal passions of a strong
man." (
This chapter's brief description of cerebral
architecture, and the functional relationships of components, is part of the
accepted neuropsychology literature. Every normal person's brain functions this
way. If the brain was a "blank slate," as Francis Bacon initially
suggested, and philosopher John Locke systematically expounded, then how
amazing it would be for the blank slate to form itself into the same well‑defined
areas, with corresponding functions, in all people ‑ regardless of their
individual upbringing and environmental experiences! This old idea is best
forgotten. Even Bacon and Locke would probably disown the outdated notion if
they were alive today and could know about recent neuropsychology findings.
