GENETIC
ENSLAVEMENT:
A CALL TO ARMS FOR INDIVIDUAL LIBERATION
Third Edition
Bruce L. Gary
Reductionist Publications,
d/b/a
5320 E. Calle Manzana
Hereford, AZ 85615
_________________________________________________________________________________________________________________________________
Published by Reductionist
Publications, d/b/a
5320 E. Calle Manzana
Hereford, AZ 85615
Copyright 2008 by
Bruce L. Gary
All rights reserved except for brief passages quoted
in a review. No part of this book may be reproduced,
stored in a retrieval system, or transmitted in any form and by any means:
electronic, mechanical, photocopying, recording or otherwise without express
prior permission from the publisher. Requests for
usage permission or additional information should be addressed to: BLG Publishing, 5320 E. Calle Manzana; Hereford,
AZ 85615.
Third Edition:
2008 September 12
Printed by Fidlar-Doubleday,
Kalamazoo, MI; USA
ISBN 978-0-9798446-0-7
_________________________________________________________________________________
Books
by Bruce L. Gary
ESSAYS
FROM ANOTHER PARADIGM, 1992, 1993 (Abridged Edition)
GENETIC
ENSLAVEMENT:
A
CALL TO ARMS FOR INDIVIDUAL LIBERATION, 2004, 2006, 2008 (this 3rd edition)
THE
MAKING OF A MISANTHROPE: BOOK 1, AN AUTOBIOGRAPHY, 2005
A
MISANTHROPE’S HOLIDAY:
VIGNETTES AND STORIES, 2007
EXOPLANET
OBSERVING FOR AMATEURS, 2007
QUOTES
FOR MISANTHROPES: MOCKING HOMO HYPOCRITUS, 2007
THE
MAKING OF A MISANTHROPE: BOOK 2, MIDNIGHT
THOUGHTS (2008)
_____________________________________________________________________________________________________________

"The topic for today
is: What is reality?"
___________________________________________________________________________________________________________________
“Men value
women because they can make babies. Women value men because they can support
and protect a family. The genes value both because their enslavement offers
a prospect for genetic immortality.” Bruce L. Gary
"So free
we seem, so fettered fast we are." Robert Browning,
Andrea del Sarto, 1855
Do you know
what the real question for a thinker is? The real question is: How much
truth can you stand?" Spoken by Nietzsche
character in When Nietzsche
Wept, by Irvin D. Yalom,
1992
"When God
is at last dead for Man, when the last gleam of light is extinguished and
he is surrounded by the impenetrable darkness of an uncaring universe that
exists for no purpose, then at last Man will know that he is alone and must
create his own values to live by." Nietzsche (altered
quotation)
“It’s a privilege to have been born and to live on this planet for a few
decades.” Richard Dawkins, in a debate June 2007
_____________________________________________________________________________________________________
C O N T E N T S
Prologue
Introduction
Outlaw Genes in 1962, Uncrossed
Paths in 1963,
Book Overview
1
Reductionism
Universe Rigid, F= ma, No Room for Spirit
Forces,
Dreiser's
"No Why, Only How"
2 Spiritual Heritage
Primitives Need Spirits, We Must Resist
the Backwards Pull
3 Genetics Tutorial ‑ Part I
Review of Earth Life, Competition is Between
Genes, Which
Genes
Compete, Gene Interaction Effects, Trade‑Offs
and
Compromises, Individual Welfare Irrelevant,
Inclusive
Fitness
4 Genetics Tutorial ‑ Part
II
Pre‑adaptation,
Species‑Shaping Forces, How Many Genes
Compete, Pace of
Evolution, Unintended Deleterious
Effects, Dangers
of Fast Evolution, Lag and
Regression, Mutational
Load, Reverse Evolution,
Pleiotropy and Polygenes
5 Genetics Tutorial ‑ Part III
Remote Sensing
Metaphor
6 Evolution Concepts and Humans
GEP, Men Bear Greater Burden of Selective Forces,
Takeover
Infanticidal Males, Monogamy and Cuckolding,
Men
and Women Shape Each Other, Birth Order,
Duality
of Morality, Emotions Control the Rational,
Consciousness
7 Brain Anatomy and Function
Vertical organization, Cerebral Lobes,
Function, Laterality
8 The Brain's Role in Evolution
Prefrontal is Recent, Modules and Genes,
Competing
Modules,
Result‑Driven Thinking, Niches, Individual
Ontogeny
& Species Phylogeny
9 Artisans Set the Stage for Civilizations - Part
I
Tool Making Artisans go Full‑Time, New
Artisan Niches
10 Artisans
Set the Stage for Civilizations - Part
II
Co‑Evolution of Niches and Genes, LB‑Driven
Rise
of Civilizations
11 Lessons from Sailing Ships
Co-evolution of
genes for Altruism/Selfishness and
Intolerance/Tolerance (Group Selection Theory)
12 Levels
of Selection, Rise and Fall of Civilizations
Gene Selection, Group Selection and Individual
Selection,
New Measure for Strength of Selective
Forces,
Rise and Fall of Civilizations, Oscillations
as a Transitional
Mode
13 The
Origins of Two Cultures ‑ Part I
Evolution of a
New Left Brain, "Resentful" Right Brains,
LB/RB conflicts
14 The
Conflicts of Two Cultures ‑ Part II
Example Newspaper Articles,
Example Books, Eastern
Thought,
Fiction and Art, Spiritual Scientists
15 Factors
Influencing Fate of Civilizations – Part I
Natural Catastrophes, Group Selection Theories
16 Factors
Influencing Fate of Civilizations – Part II
Producers/Parasites,
RB/LB Conflicts, Sexual Selection
17 Factors
Influencing Fate of Civilizations – Part III
Troubadours,
Women Speed Civilization’s Fall
18 Factors
Influencing Fate of Civilizations – Part IV
Turning
Inward, Mutation Load (dysgenia)
19 Factors Influencing Fate
of Civilizations – Part V
Fascism Causing Collapse of American Empire
20 Dating the Demise of Humanity
New Time Scale for Humanity, Doomsday Argument
and
Anthropic
Principle, Probabilities of Population Collapse
21 A Global Civilization Crash Scenario
Tribalism's Starring Role; Communism &
Fascism
as
Twin Gene‑Driven Enemies of Artisan‑Created
Civilization,
Genetic Entrenchment and Culturgens,
Conformism,
RB's Revengeful Victory Over LB
22 Living Wisely ‑ Seeking Positives
Mount
Cognoscenti,
Life Dilemmas, Eschewing the
Crowd,
Activity Categories, Emphasize Positives,
Brief
Encounters
23 A Call to Arms ‑ Identifying Outlaw Genes
Prospects
for Replacing Gonad Man After the Crash,
Genetic
Pitfalls
24 Utopias
Isolated Communities, Cognoscenti Societies,
Platonic
Aestheticism
25 Repudiation of the Foregoing
Ultimate Meaninglessness of Everything,
A Hierarchy
for
Dealing With Reality, Existentialism
26 A Free Man's Worship
Annotated
Version of a Bertrand Russell Essay
Your Odyssey
Appendix A: Reductionism
Appendix B: Human
Virus Examples
Appendix C: Remote
Sensing Analogy
Appendix D: World
Population Equations
Appendix E: More
Repudiation of the Foregoing
References
Index for Authors
Index for Words
_______________________________________________________________________________________________
.
PROLOGUE
"Generally
speaking, it is quite right if great things ‑ things of much sense for men
of rare sense ‑ are expressed but briefly and (hence) darkly, so that barren
minds will declare it to be nonsense, rather than translate it into a nonsense
that they can comprehend. For mean, vulgar minds have an ugly facility for
seeing in the profoundest and most pregnant utterance only their own everyday
opinion." Jean Paul, as quoted
by Friedrich Nietzsche, Philosophy in the Tragic Age of the Greeks,
1872.
Dear reader,
you normaloid idiot!
Well, maybe you
deserve an explanation for that greeting.
A perceptive alien
visitor to Earth might report home that humans are the dumbest and most
despicable creatures on the planet!
At least the other
animals don’t claim to know things which, in fact, are absurd nonsense.
Only humans believe in such imaginary things as heaven, hell, guardian angels,
telepathy and all kinds of gods. Only humans maintain that the world was
created by some imagined godly entity just for them and that this God continues
to watch everything and tests humans so that He may reward or punish them
in accordance with how pleased He is by their behavior. Only humans believe
that they are so different from non-living things that their “consciousness”
exempts them from the laws of physics. But the most incriminating human trait
is that homo sapiens is the only species that has itself
for its most dangerous enemy, and a revealing irony is that most killing is
done on behalf of this thing they call “religion.”
Human conceit and
imagination is so poor that people cannot imagine themselves as automatons
that are assembled by genes. Even those few humans who do accept that they
were assembled by genes seem unable to imagine that these genes have achieved
longevity in the species gene pool by assembling automatons that serve
those very genes instead of the individual. This saves them from the indignity
of realizing that they are foolish slaves to tiny lifeless molecules that
use them for aimless ends.
The humans, these
aliens might conclude, are hopeless!
So now, dear reader,
we must have a delicate conversation about you in relation to this book.
If you are like that clueless 99% of humans, those I call “normaloids,” then
let me suggest that you abandon this book and resume your pathetic, unthinking
life! You may do so now! Please do so now!
Are you still reading?
Are you a normaloid pretending to be one of that 1% of thinking humans?
I give you one last chance to feel the guilt of reading something not meant
for you.
Cognoscenti
The following was written for the diminishing numbers
of “the cognoscenti.” And to the cognoscenti who may be holding this book,
I apologize for writing things that are inherently self-evident. You may
have already thought of them yourself, and gone beyond my modest collection
of thoughts. But if, by chance, you have not already discovered the self-evident
ideas in this book then I hope you enjoy the following.
Reductionism
and Hypocrisy
I'm a robot! So
are you! This book views people as robots assembled by genes for the "purpose"
of serving them by behaving in ways that have led to genetic prosperity in
the ancestral environment. Only this “reductionist” viewpoint provides insight
into the many bizarre aspects of human nature.
Every thinking person
should be disappointed in humanity! Indeed, every thinking person should
become a “misanthrope.” In youth it is easy to idealize human nature, to
believe what people say about themselves. Later, perhaps in the teen years,
human hypocrisy is discovered. The so-called “pursuit of Truth” becomes a
hollow promise. Adults who continue to believe in childish notions of human
nature look foolish.
I’m more disappointed
than bitter. I can say that with each year's accumulation of disappointment
in human nature my interest in writing this book wanes. Among the plethora
of book publications there are only a handful for the reader who knows how
to think. Even most of those intended for serious reading are fundamentally
flawed. Why, I keep asking, are so many people incapable
of thinking!
Alas, there is an
explanation; an explanation, indeed, for all the flaws in human nature! We
are the way we are because the genes have constructed us this way because
it serves them!
The genes that assemble
us were survivors in the "ancestral environment" (AE). Not only did they
make fools of us in the AE, but in the modern environment our inherited tendencies
make new fools of us in ways that were not even anticipated by the genes.
Anyone who occasionally
glimpses humans this way has the opportunity of choosing a path leading to
a belief that humans are victims of genetic enslavement.
Life takes on new meaning for the person who then wishes for liberation from
that enslavement. This book is dedicated to that rare person already on such
a journey of liberation.
The
mind is a terrible thing to trust
Humans are severely
handicapped at comprehending such things as sub‑atomic strings vibrating
in 11 dimensions, a universe that will expand forever and cause all matter
to "evaporate" in 10100 seconds, or even the everyday experience
of seeing a commercial jet airplane that appears to be 35 degrees ahead of
where the sound is coming from. The list of things we are ill-equipped to
understand is immense!
We cannot readily
understand these things because they never affected the survival of our ancestor’s
genes. How many more aspects of our world are inherently elusive because
they never mattered to genetic survival? Or worse, how many things are hidden
from us because they belong to a category of knowledge that would have adversely
affected the survival of the genes our ancestors carried, even though this
insight might have enlightened the individual?
The layman seems
stubbornly committed to the belief that our minds can be trusted to have
an intuitive understanding of all things. Both the layman and professional
alike will instinctively object to any suggestion that our genes construct
brains that "intentionally" handicap our ability to comprehend the way the
genes have enslaved us. To put it bluntly, I am suggesting that our minds
are designed to steer us away from Truth when alternative false beliefs safeguard
genetic enslavement of the individual, even when this blinded vision diminishes
individual well‑being.
Humanities
versus Physical Sciences
Don't expect humility
from humans. Just as every serious thinker must become exasperated with others,
so should he become exasperated with himself (I use "him" instead of "him/her").
Even within the physical sciences, where I earned a living for 43 years,
it is necessary to consciously maintain vigilance against well‑meaning, intruding
intuitions. Imagine how difficult the task must be within the humanities,
which are blatantly undisciplined compared to the physical sciences. Physical
scientists deal with quantifiable predictions which can be tested by observations.
In the humanities, on the other hand, practitioners seem more concerned
with loyalty to charismatic leaders, and their beliefs, than to the pursuit
of objective truth. Imagine, then, how easily investigations in the humanities
can go astray.
And gone astray
they have! The long endeavor to understand "human nature" has had more false
leads from well‑meaning professionals with social agendas than probably any
other field. For example, some people contend that "human nature" doesn't
exist, believing instead that our minds are "blank slates" at birth, ready
to be written upon for the creation of whatever mental structures conform
to the external world. Others state that “human races” don’t exist, yet insist
on affirmative action preferences for
non-existent minority races. Such beliefs are congenial to
those who secretly wish to fiddle with the social environment for the purpose
of correcting social injustices. Marxist minds are naturally attracted to
the humanities, and have tried for nearly a century to hijack anthropology
and distort it for their purposes.
In spite of the
odds against progress, and in spite of energetic people who seem bent on
leading others astray, there are achievements to be proud of in the study
of human nature. Anthropology and psychology may have a sordid record of undisciplined
meddling by people with political agendas, yet uphill progress in these
fields has surely occurred.
Academic
Quarrels
I recognize that
most readers will object to this misanthropic portrayal of human nature and
my cynical description of "human behavioral scientists." They may be inclined
to agree with some of it, but they will quibble with specifics, or insist
on different ways of approaching the subject. Just as tribes need to fission
when they become too big, major subject areas within academe need to splinter
to form "schools of thought" that go their separate ways by maintaining
petty quarrels. For example, evolutionary psychologists complain about sociobiologists
not having the proper "nuance" concerning adaptation versus optimization,
and they use this minor complaint to build a wall of separation when as
a practical matter the two fields are essentially one.
I am mindful of
the need for petty carping by academics, or the inevitability of it, but
I deplore the loss of vision that it inflicts upon those caught‑up in it.
Sometimes a professional becomes so involved with argument over petty differences,
and concern over whose grant request will be funded, that he forgets to stand
back from day‑to‑day controversies in his field to see it in the larger perspective.
The preoccupation with professional details may render the professional
practitioner blind to bigger visions that can only be seen from a distance.
An outsider, looking in, will occasionally be worth listening to, for he
brings with him that distant "big picture" perspective. I claim to bring
a "big picture" perspective to the subject of sociobiology, and this should
interest the serious lay reader as well as the professional sociobiologist.
This book asks a
lot from the reader without a background in sociobiology, and I realize that
few, if any, will read it through. The professional sociobiologist will readily
understand most of my message, but he will be troubled by the fact that
he does not recall reading other articles by me in sociobiology journals.
The lay reader will not be bothered that my publications are in a totally
unrelated field, but he will find much of the material unfamiliar and will
be repelled by it.
I will not be disappointed
if neither the sociobiologist nor the lay person reads what follows. My life-long
romp in the realm of ideas, and my writing of essays that appear in this
book, has been more fun than what I imagine it would be like to have positive
reader feedback or book sales. Indeed, as of this Second Edition writing
(2006 January) fewer than a dozen of the first edition have been sold.
When
I’m optimistic I recall Henri Beyle (Stendhal), who believed that his writings
would escape notice until a century after his death. His forecast was amazingly
accurate. Such a fate could in theory happen to this little book, but I now
realize that the process of creating it was reward enough. I had more fun
writing it than any reader could possibly experience in its reading. Like
any creation, this book was written for the author.
.
─────────────────────────────────
INTRODUCTION
─────────────────────────────────
BEGINNINGS
OF AN IDEA AND BOOK OVERVIEW
Washington, DC in 1962 was an
exciting place. President Jack Kennedy created a “Camelot” aura that fed
hope for unbounded progress. But the Cuban Missile Crisis brought a sobering
chill to the country, especially to residents of Washington, DC. On my way to work
I'd look north at the Capitol Building and wonder if it
would be blown‑up by a Soviet missile while I was looking at it.
My first job after
college was at the U.S. Naval Research Laboratory, where I worked as a radio
astronomer specializing in Jupiter's radiation belts. Freed of time‑consuming
college coursework, I was able to broaden my reading. A few years earlier,
the double‑helix structure of DNA had been discovered. Perhaps stimulated
by this, or maybe from the sheer momentum of a childhood fascination with
the way genes influence behavior, I stumbled upon a thought which I now believe
is the second‑most profound one of the 20th Century: “outlaw genes.”
1963 Identification
of Outlaw Genes
On February 23, 1963 I was imagining
the possibility of categorizing gene mutations as either promoting or subtracting
from their ability to survive into the future and I needed terminology for
this gene attribute. "Gene Survival Value" came to mind. Given a sufficiently
well thought out measurement protocol any gene could theoretically be placed
on a GSV spectrum, with endpoints labeled PGSV and NGSV - standing for "positive
GSV" and "negative GSV." (I recall being dissatisfied with such awkward terms).
At about the same time I was also struggling to devise theoretical concepts
that might guide an individual in choosing a "rewarding life path," as ill‑defined
as such a concept can be in youth. Longevity was one factor, so given the
GSV example I invented ISV, for Individual Survival Value. The ISV extremes,
of course, were PISV and NISV. At this critical juncture, it seemed right
to draw an X‑Y coordinate system, representing GSV and ISV. (In retrospect,
"individual well‑being” would have been a better parameter to adopt than
Individual Survival Value.) The figure on the next page is a rendition of
this scatter diagram.
In theory, any gene
could be "placed" in such a diagram (I hadn't encountered the concept of
polygenes or pleiotropy at that time, to be discussed in a later chapter).
I imagined genes for this and that, and placed them in the diagram. I recall
thinking that there had to be more dots in the upper‑right quadrant, corresponding
to PGSV/PISV.
I realized that
there shouldn't be many dots in the opposite corner since NGSV/NISV mutations
should quickly disappear. Likewise, there shouldn't be many dots in the upper‑left
NGSV/PISV quadrant, though wouldn't it be nice if genes flourished when
they promoted individual happiness regardless of the cost to themselves.
But it was the lower‑right corner that awaited me with a surprise! Gene mutations
of this type would "by definition" flourish while "punishing" the individual
carrying them! And nothing could be done about it, short of replacing the
forces of natural selection with artificially created ones. This gene category
has fascinated me ever since!
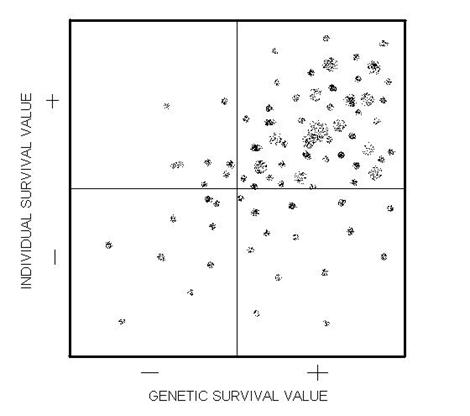
Figure 1.1 An X‑Y matrix
of "genetic survival value" and "individual survival value" with hypothetical
markings of the locus of individual genes (as conceived in 1962).
Why hadn't I read
about such genes? Surely others knew about the inherent conflict between
the individual and some of the genes within! I looked forward to someday reading
about these "outlaw genes," and the philosophical dilemmas they posed. I
stashed these original diagrams and writings on the matter in a file, which
remained closed for decades. Nevertheless, I did not forget about these genes
and during the past four decades I have written about the subject in my
spare time.
Coincidences
In the Fall of 1963 I enrolled at the University of California at Berkeley for graduate studies
in astronomy. As the prospect of taking required courses on such topics as
stellar spectroscopy sunk in, I realized that my career path had taken a
wrong turn, of sorts, since my heart was with the humanities. I managed to
add classes in psychology and anthropology as a consolation for the dry astronomy
stuff. (I quit before semester's end, and have been
gainfully employed in the physical sciences ever since.)
Although coincidences
can shape lives, more often they don't. While I was at Berkeley a little‑known
biologist, George C. Williams, was using the school library to write a
manuscript that would be published in 1966 as Adaptation
and Natural Selection: A Critique of Some Current
Evolutionary Thought. He was making a case for the view that selection
forces work at the level of the genes, not the individual (and definitely
not the species). Although this perspective was inherent in my thinking I
failed at the time to grasp its novelty. I assumed that somewhere in the humanities
was a field in which everyone believed this. Of course I was wrong, for Williams
was engaged in creating such a field.
In this same year,
1963, William D. Hamilton prepared manuscripts describing "inclusive fitness"
(Hamilton, 1964a,b), which is an essential part of understanding how
gene competition drives evolution. The work of both Hamilton and Williams
were essential footings, one decade later, for Edward O. Wilson's milestone
book Sociobiology: The
New Synthesis (Wilson, 1975). In my opinion,
sociobiology is the most important idea of the 20th Century.
I sometimes wonder
how my life's path might have differed if I had met Williams at Berkeley in 1963. A conversation
with him could have clarified for me the emerging nature of the new field,
and the opportunity for a role that I might have played in that emergence.
Although the field was closer to my heart than astronomy, I never ran into
G. C. Williams, and I never realized that he was helping to give birth to
"my" field.
Overlooked Idea
Even now, four decades
later, no one has written clearly about the mischievous genes (to my knowledge).
The Selfish Gene, by Richard Dawkins (1976),
comes close; but it never explicitly states that genes "enslave" the individual
for their selfish advancement while harming the enslaved individual. Mean Genes (Burnham and Phelan, 2000) comes
even closer, but its emphasis is on practical steps for resisting self‑defeating
behaviors rather than the theoretical origins of the genes responsible for
those behavioral predispositions.
Why is there such
a paucity of discussion about the philosophical implications of such a profound
flaw in our origins and present nature? Why have the professional anthropologists,
philosophers and others been so slow to address a subject that captured my
unwavering attention 40 years ago, when I was fresh out of college and struggling
to establish a career in an unrelated field? Sociobiologists have written
about conflicts between competing gene alleles carried by individuals of
various relatedness (Hamilton, 1964a,b), between parents and offspring (Trivers, 1974),
and between siblings (Sulloway, 1996), but not between the individual and
his genes! If any field has a mandate to ask the questions I stumbled upon
in 1962 it is the new field of sociobiology!
If my idea has merit
then sociobiologists have simply overlooked an obvious “next step” in the
unfolding of implications for the basic tenet of the field. The history of
science has many examples of simple yet profound new ideas being overlooked
by the professionals. Every idea has many discoverers, and probably most
of them only half realize the import of their discovery. The oft‑discovered
idea remains out of the public domain until it is grasped by someone having
the energy to push it into the mainstream.
Some of the genes
within us are enemies of the individual, in the same sense that outlaws are
the enemies of a society. This thought should challenge the thinking of
every sentient being. The discipline of philosophy should be resurrected,
and restructured along sociobiological precepts. If this is ever done the
new field would have as its major philosophical dilemma the following question:
"What
should an individual do with the mental pull toward behaviors that are harmful
to individual welfare, yet which are present because they favor the survival
of the genes that create brain circuits predisposing the individual to those
behaviors?"
In other words,
should the individual succumb to instincts unthinkingly, given that the gene‑contrived
emotional payoffs may jeopardize individual safety and well‑being? Or, should the individual be wary of instincts and thoughts
that come easily and forfeit the emotional rewards and ease of living in
order to more surely live another day - to face the same dilemma? Should
some compromise be chosen? How can any thinking person
fail to be moved by these thoughts?
Overview of This
Book
In writing this
book I have wrestled with the desire to proceed directly to the matters of
outlaw genes, and how an individual might deal with them. But every time
I returned to the position that a proper understanding of the individual's
dilemma requires a large amount of groundwork. For example, how can I celebrate
the artisan way of life without first describing why the genes created the
artisan?
In the first edition
of this book I included the many groundwork chapters in their entirety before
the culminating chapters. The first person to read the book (Dr. M. J. Mahoney)
stated that “Once I hit Levels of Selection [Chapter 11] I couldn't
put the book down.” That’s when I realized that I had violated the first
principle of writing, which is to “quickly engage the reader before you lose
them.” In this edition I have shortened the groundwork chapters by moving
most of that material to appendices. The groundwork chapters have become a
primer for the paradigm that leads inevitably to the positions of the main
message of this book.
The remainder of
this introduction is a précis for the book chapters.
There is no guiding
hand in evolution; the natural process of the genes acting on their own behalf
leads to individuals who are mere "agents" for these genes. This is the
perspective of "sociobiology," also called "evolutionary psychology," and
presented most effectively for the general public by Richard Dawkins in
The Selfish Gene (1976). To understand the
"blindness" of evolution one must first understand that the universe is just
a "mechanism," that every phenomenon reduces to the action of blind forces
of physics acting upon dumb particles. This outlook is called "reductionism,"
and is the subject of Chapter 1.
Lest the reader
surmise that this book is about the physics of life, I attempt an impassioned
appeal, in Chapter 2, for an embrace of modern man's scientific approach
to understanding life, and a rejection of the primitive backwards pull that
captures most unwary thinkers. This appeal provides a foretaste of the spicy
sting of chapters found in the second half of the book.
Since genes are
such an essential player in everything, I found it necessary to include tutorial
chapters on genetics. The first of these genetics tutorials, Chapter 3,
presents general properties of genes, such as how they compete and cooperate
with each other, and have no concern for individual welfare beyond what
serves them. The second genetics tutorial, Chapter 4, explores some
subtle properties of genes that will be needed by later chapters. For example,
since in every new environment some genes will fare better than others,
it is useful to think of genes as being "pre‑adapted" and "pre‑maladapted"
to novel environments. This will be an important concept in considering artisan
niches in the modern world.
Chapter 5 is not necessary
for the development of the book’s theme, but for those who understand it
the chapter will provide a deeper insight into the mathematics of pre-adaptation
and pre-maladaptation.
Chapter 6 pulls together
some of the genetics ideas and applies them to human evolution. Certain insights
are needed for a person to intelligently deal with emotions that control
or attempt to discredit intellect. For example, how can a person handle jealousy
without understanding cuckoldry?
Chapters 7 and 8 are devoted to the
brain. The most recent advance in the evolution of the human brain is the
refashioning of the left prefrontal cortex. It is important to view the brain
as an organ designed by the genes to aid in gene survival. Rationality is
a new and potentially dangerous tool created by the genes, and it must be
kept under the control of "mental blinders" to assure that the agendas of
other genes are not thwarted. Competing brain modules, cognitive dissonance,
and self‑deception, are just a few concepts that any sentient must know about
when navigating a path through life's treacherous shoals.
In Chapter 9
I write about the first artisan, whose precarious role as a full‑time tool
and weapon maker may have begun 60,000 years ago. When the climate finally
warmed 11,600 years ago at the start of our present "interglacial," called
the Holocene, the small number of existing artisan roles served as a model
for an explosion of new ones. The new artisans made high‑density populations
possible and eventually led to the creation of civilizations (Chapter 10). Since I will celebrate the artisan way of life
it is necessary to understand how it came into existence and why others in
society are likely to view it warily. I will outline a theory for "anti‑intellectualism"
and suggest that it may play a role in a civilization's decline.
Group selection
still attracts controversy, and I use it argue that tribal warfare led to
ever‑larger tribes, which required that its membership be ever‑more subservient
to "tribal requirements" since the entire tribal membership had a shared
destiny. But, as I argue in Chapter 11, when group selective forces
were at their maximum during the Holocene, something new happened that heralded
the first‑ever "individual selection" dynamic. The artisans assumed a leadership
role in molding culture, governance, and opening opportunities for individual
expression of creative and productive labors that led to a state that we
now call "civilization."
But a civilization
is vulnerable to outside attack by societies that remain uncivilized, that
foster religious fanaticism. These stay‑behind societies harbor resentment
of the material wealth of the civilized society, and instead of achieving
wealth for themselves by surrendering their group‑serving grip on the individual,
they instead mobilize the individual to discredit their rich neighbor and
declare cultural warfare on them. Religious zeal serves these super-tribes
by fostering fanatical, suicidal attacks on those societies that respect
the individual. But since individuals in the civilized society think first
of themselves, the civilization's defense is half‑hearted and ultimately ineffective.
It is inevitable
that civilizations arise with an ambivalent self‑hatred. This is because
people whose thinking style is overly influenced by their "primitive" right
brain are naturally resentful of the world created by those new left‑brain
artisans. The new world order favors the left‑brained artisan (engineer, scientist
and other rational thinkers) and relegates to some vague periphery the contributions
that can be made by the old‑style people. Thus, every civilization should
have "two cultures" that are in conflict, and this is treated in Chapters
12 and 13.
Chapter 14 begins to address
the matter of what factors might contribute to the decline and fall of civilizations.
One theory invokes a back‑and‑forth dominance of artisan "producers" versus
opportunistic "parasites." Another suggests that the two cultures war, or
the “War of the Brain Halves,” is eventually won by those who succumb to
the primitive pull. Gingerly, I also suggest that dysgenia might undermine
our genetic vigor and sap societal energies.
I discovered the
Anthropic Principle (and learned that it had been written about and published
obscurely a few years before my discovery of it). I use this idea to predict
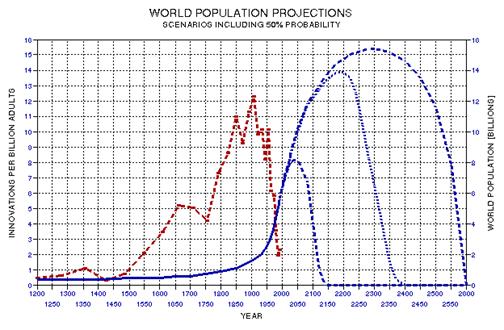
the approximate date range for a significant crash in the human population. In the process of calculating this horrific event, I show
that the rate of technological innovations exhibits a trace over time that
foretells population patterns. From this analysis it appears that we are
now in the second major "rise and fall" pattern of innovation rate and population,
the latter pattern being displaced a few centuries after the first. This
is described in Chapter 15.
I attempt to survey
some possible population crash scenarios in Chapter 16. However, I
conclude that the future is so difficult to predict that it is prudent to
only present possibilities.
In Chapter 17
I begin my "call to arms" for individuals to emancipate themselves from the
genetic grip. All previous chapters are preamble to this one and those that
follow. My appeal must be qualified by some nitty‑gritty facts of genetics,
such as pleiotropy and polygenes. Nevertheless, I present a litany of "genetic
pitfalls" that any emancipated person should wish to avoid.
Because any reader
will expect a book such as this to give specific suggestions for how to use
insight to live wisely, I feel obligated to present in Chapter
18 my feeble attempt to address the subject. It is an attempt to describe
ways that an individual may live wisely in a world wracked with defects caused
by outlaw genes. Some genes are our enemy because they lead to dysfunctional
human societies, while other genes are our enemy because they lead us as
individuals to want the wrong things. The individual's task is to liberate
oneself from the genes, and choose wisely. The IQ form of intelligence allows
insight, and this insight must be placed into the service of an enlightened
"emotional intelligence" to arrive at new personal values to live by. The
questing person will understand the wisdom in the saying, which applies to
the unthinking person: "If you get what you want, you deserve what you get."
However, I readily acknowledge that my attempt to realize this chapter's
goal is feeble, and the reasons for this are developed at the end of the book.
Chapter 19 follows naturally
from the previous chapter, since an individual who wishes to pursue an individual‑emancipated
life must do so within the constraints of living in a society where individual
liberation is difficult. When a sufficient number of people awaken to their
enslaved condition, thoughts may turn to a way for them to coalesce in
a shared search for a winning place. I describe utopias and prospects for
isolated enclaves as a path toward a stable community where individual liberation
may be sought. However, I warn that the world is becoming too "small" for
enclaves to remain safe from meddlesome outsiders. Since the door of feasibility
for creating isolated space communities has shut, and since the earth is
already "too small" for self‑sustaining communities to remain secret, there
are no feasible refuges for utopias. I conclude that today's world will not
tolerate the formation of an enlightened society of liberated individuals,
and that those who might wish to live in such a society must be content with
learning how to live a good life as individuals with secret dreams while being
surrounded by an ever‑increasing number of primitive hoi poloi.
The "society of the cognoscenti" will remain dispersed, and may only occasionally
recognize each other during normal encounters.
Chapter 20 is supposed to
be a surprise, but the subtitle sort of gives it away: Repudiation of the
Foregoing. I will say no more.
Chapter 21 is an annotated
version of Bertrand Russell's essay, “A Free Man's Worship.” It is an excellent
example of how a liberated person thinks, and I use it to illustrate the
point of the preceding chapter. Namely, once a person is liberated from genetic
enslavement and free to choose values to live by that are compatible with
the cognoscenti's insights, an aesthetic and poetic
attitude toward "existence" can be achieved. The existentialist need not
be a sourpuss, nor must he become a passive esthete. The thoughtful existentialist
may end up a compassionate humanist with a lust for existence!
So now dear reader,
if you exist, do take the following speculations with a light heart; hopefully
your thoughts will be led in directions that are as congenial to your inherited
ways of thinking as the following are to mine.
─────────────────────────────────
CHAPTER
1
─────────────────────────────────
REDUCTIONISM
An
intellect which at any given moment knew all the forces that animate Nature
and the mutual positions of the beings that comprise it, if this intellect
were vast enough to submit its data to analysis, could condense into a single
formula the movement of the greatest bodies of the universe and that of
the lightest atom: for such an intellect nothing could be uncertain; and
the future just like the past would be present before its eyes. Pierre Simon Laplace,
Philosophical Essay on Probabilities
(1814).
The
highest object at which the natural sciences are constrained to aim, but
which they will never reach, is the determination of the forces which are
present in nature, and of the state of matter at any given moment ‑ in one
word, the reduction of all the phenomena of nature to mechanics." Gustav Robert
Kirchhoff, 1865.
The idea that all
movement could in theory be reduced to simple laws of nature was first recorded
for posterity by Democritus of Abdera in 4th Century BC Greece. This reductionist
outlook was also expressed by the Roman philosopher Lucretius during the
1st Century BC. After the Dark Ages, when Greek and Roman ideas
lost favor, disillusionment with religion grew among thinkers and they became
curious about the ideas of those ancient challengers of piety and ritual.
Isaac Newton wrote about the forces of nature as a basis for understanding
movement during the 17th Century. The Philosophes
of 18th Century France restored rationalism as the ultimate guide
to Truth. Scientific discoveries continued to provide support to the reductionist
notion, as shown by leading 19th Century scientists such as Laplace
and Kirchhoff.
During the early
20th Century the reductionist paradigm came under challenge by
Quantum Physics. The new physics does not claim to require divine intervention
or primitive spirits, but it does appear to require randomness for events
at physical scales the size of the atom and smaller. The so-called Newtonian
physics is correct as far as it goes, but it cannot explain a category of
physical events associated with the atom and its interaction with light.
At some future time it may be possible to portray quantum physics with the
same deterministic quality as Newtonian physics, but for now it is prudent
to assume that some events are subject to random outcomes. The strict form
of determinism called for by Newtonian physics should be replaced by a probabilistic
form of determinism. However, both forms of determinism are reductionist
since they are based on the notion that the movements of all particles, and
their interaction with light, conform to the laws of physics. This last phrase
is key, “conform to the laws of physics,” and it is dealt with at greater
length in Appendix A.
"Reductionism" either
angers people or delights them. The entire enterprise of science is based
on reductionist tenets. Whereas all scientists practice their profession in
accordance with the reductionist paradigm, there’s a part of the brain that
is so opposed to it that ~40% of scientists claim to not believe in reductionism.
These conflicted scientists are found mostly in the humanities, where muddled
thinking is less of a handicap; within the physical sciences there is almost
universal belief in reductionism (especially among the most esteemed scientists).
The end-point of
reductionist thinking can be most easily described by the following analogy:
the universe is like a giant billiard table, in which all movements are mechanical
‑ having been set in motion by the explosive birth of the universe 13.7
billion years ago! Of course this analogy neglects quantum physics, but
nothing essential to our understanding of the evolution of life, and of
human nature, is lost by this simplification. The march of events, from one
moment to the next, is captured by this simple-minded, deterministic view.
Since this version of reductionism is essential to the rest of this book,
and since it is a fascinating subject in its own right, I devote the rest
of this chapter to a brief description of reductionism and an appendix to
its fuller treatment.
A Rigid Universe
"The Universe Rigid"
was possibly H. G. Well's most important manuscript. It languished with the
publisher, who didn't understand it, and eventually it was lost. Instead
of reconstructing it, Wells turned its central idea into a story, The Time Machine (1895).
Two dozen years
later, Albert Einstein published his Special Theory of Relativity (1920),
which expanded upon the idea, already familiar to physicists of the day,
of time as a fourth dimension. This concept was treated in "The Universe Rigid"
essay with unusual insight, especially for a non-physicist. The following
is a brief summary of it that appeared in The Time
Machine:
"...
Suppose you knew fully the position and the properties of every particle
of matter... in the universe at any particular moment of time:.. Well, that knowledge would involve the knowledge
of the condition of things at the previous moment, and at the moment before
that, and so on. If you knew and perceived the present perfectly, you would
perceive therein the whole of the past. ... Similarly, if you grasped the
whole of the present, ... you would see clearly
all the future. To an omniscient observer ... he would
see, as it were, a Rigid Universe filling space and time ‑ a Universe in
which things were always the same. He would see one sole unchanging series
of cause and effect... If ‘past' meant anything, it would mean looking in
a certain direction; while ‘future' meant looking the opposite way. From the
absolute point of view the universe is a perfectly rigid unalterable apparatus,
entirely predestinate, entirely complete and finished... time is merely a
dimension, quite analogous to the three dimensions of space."
This passage describes
the underlying principle of what is referred to in today's parlance as "reductionism." It leaves no room for spirits, mysticism or gods. Most
intellectuals use the term "reductionist" as a disparaging epithet, but I
believe that reductionism is the crowning achievement
of human thought.
Mechanical Materialism
Reductionism
is the belief that complex phenomena can be "reduced" to simpler physical
processes, which themselves can in theory be reduced to the
simplest level of physical explanation where elementary particles interact
according to the laws of physics. The ancient concept of Mechanical Materialism
captures the essence of reductionism, but relies upon the outdated concept
that at the most basic level the particles are like stones, and interact
by hitting each other like billiard balls.
Nevertheless, reductionism is the fulfillment of what Democritus and Lucretius
dreamed about, a mechanistic world‑view that as a bonus could also deliver
people from the tyranny of religion. Lucretius would agree with the statement:
"There is no need for the aid of the gods, there is not
even room for their interference.... Man's actions are no exception
to the universal law, free‑will is but a delusion." (Bailey,
1926, describing how Lucretius viewed the world).
It will be instructive
to review mechanical materialism before describing the version of reductionism
required by modern physics.
Imagine a game of
billiards photographed from above, and consider frames redisplayed in slow
motion. After the cue sends one ball into motion, the entirety of subsequent
impacts and bounces are determined. If this were not so, if the balls had
a mind of their own, or if some mysterious outside force intervened, then
consistently good players would not exist. Now imagine a very slow replay
of the motions of the billiard balls; millisecond by millisecond the movements
unfold with an undeniable inevitableness. A careful analysis would reveal
that sustained momentum and elastic collisions govern the placement and velocity
of each ball in the next millisecond.
Given two successive
"frames," an observer would know the positions and velocities of every ball,
and he could calculate their placement, velocities and future impacts for
any arbitrarily short instant later. He could thus predict the following
frame, and the process that allowed the prediction of frame 3 from frames
1 and 2 could be repeated for frames 2 and 3 to predict frame 4. And so on,
for all future frames. In this way, the observer could
predict all future movements (don’t worry about the fact that we've ignored
friction).
By a similar process
the observer could infer a previous frame from any two neighboring frames.
Thus, frames 1 and 2 could be used to predict frame 0, etc. Therefore, by
knowing any two frames, all future and past frames could be inferred. This
is the thought H. G. Wells captured with his unpublished Universe
Rigid essay.
Reductionism as
a Basis for Physics
The 19th Century
saw a de‑mystification of various science disciplines. The reshaping was
done by rationalists, building upon the legacy of the 18th Century philosophes. The rationalists firmly placed science on a footing
that has endured throughout the 20th Century. Like the machines of 19th Century
inventors, the paradigm developed by 19th Century scientists was "mechanistic."
Ernst Mach forced
metaphysics out of physics (Holton, 1993, pg. 32). Chemistry was changed
from a floundering quest for transmuting common elements to gold, into a physics‑based
understanding of atoms and molecules. Darwin displaced God from
the creation of life by presenting his theory of evolution, even though this
may have saddened him personally.
By the end of the
19th Century, when Wells began to write, the intellectual atmosphere was
congenial to ideas that reduced mysterious happenings to a juxtaposition of
commonplace physical events. Each event in isolation was conceptually simple.
It is the mere combining of many such events that cause things to appear
incomprehensible.
Reductionism is
based on a concept taught in college Physics 101. I remember well that without
fanfare the physics instructor stated that there are only four forces in
nature (gravity, electro‑magnetism, the nuclear force and the weak force),
and that these forces act upon a finite number of particles that are pulled
this way and that by the summation of all forces acting upon each particle.
In laboratory experiments where the number of relevant forces can be confined
to only 1 or 2, motions are observed to be governed by a simple law: F = m•a, or "force equals mass times acceleration"
("acceleration" is the rate of change of "velocity vector"). It is easier to understand this law of nature by rewriting
it in the form: a = F/m, which states that
a particle's acceleration is proportional to the sum of forces acting on it
divided by the particle's mass. Mathematically, a and F are vectors, which is why these
symbols are written in "bold" typeface, and "m" is a scalar (no orientation
is involved); thus, the equation a = F/m keeps
track of the 3‑dimensional orientation of forces and accelerations. Since
forces can originate from many sources, they must be added together to yield
one net force.
At every instant
a particle is responding to just one net force. It responds by accelerating
in the direction of that force (which has a magnitude and direction). The particle's velocity vector changes
due to its acceleration. Since the time history
of a particle's velocity specifies where it goes, the particle's "behavior"
is completely determined by the forces acting upon it. This description is
called Newtonian physics, and it reigned supreme throughout the 19th
Century.
Quantum Physics
During the late
19th Century a disturbing number of laboratory measurements were
made that defied explanation using Newtonian physics. Radioactivity was a
puzzle, for it seemed that atoms of certain (radioactive) elements would
spontaneously, and at random, emit a particle. There was also the puzzle of
atoms absorbing and emitting light at only specific wavelengths, producing
a unique spectral pattern for each atomic element. Newtonian physics had no
way to accommodate these and other puzzling phenomena.
Quantum physics
was developed in response to these puzzling measurements, all of which were
related to mysterious phenomena inside the atom. The new physics expanded
upon the idea that everyday objects were constructed from electrons orbiting
a nucleus composed of protons and neutrons (now known to be constructed from
12 elementary building blocks of matter). It was
proposed that electrons could be thought of as a wave, with a wavelength
such that the only permitted orbital circumferences around a nucleus were
those with an integer number of wavelengths. Changes in an electron’s orbit
involved changes in energy, so if an electron moved to a higher energy orbit
(farther from the nucleus and larger in circumference) it must absorb energy
from somewhere (such as a photon of light) that had an energy corresponding
exactly to the difference in the electron’s energy in the two orbits – hence
the quantization of spectral absorption features for each atomic element.
As quantum
physics developed to explain more laboratory experiments related to the atom,
the theory became weirder and weirder. Quantum mechanics (QM) was developed
to deal with particles, and quantum field theory was developed to explain
radiation and its interaction with particles. Quantum physics has been
described as inherently probabilistic, or indeterminate, and has been characterized
as having so much "quantum weirdness" that our minds are intuitively unprepared
to comprehend it. Quantum physics “works” in the sense that it gives a better
account than any other theory for atomic scale physical phenomena. Contrary
to popular belief, it does not discredit Newtonian physics, which is still
valid for large scale phenomena; rather, it is more correct to say that
quantum physics supplements Newtonian physics. Almost
every physical situation can be easily identified as requiring one or the
other embodiment of physical law.
It now seems
that two of the aforementioned four forces can be "unified" (the weak and
the electromagnetic). One of the main goals of physics today is to create
a “unified” theory that incorporates all the explanatory power of the four
forces plus the weird but useful explanatory power of quantum physics.
One of the
most counter-intuitive properties of quantum physics is the notion that events
are not strictly determined but are only probable, and that particles are
not tiny things at a specific location but are probability functions in
3-dimensional space. When a particle moves the probability
function describing its location moves. In the laboratory it is impossible
to measure a particle’s position without changing its velocity; and it is
similarly impossible to measure a particle’s velocity without changing
its position. The Heisenberg Uncertainty Principle quantifies the partitioning
of position and velocity uncertainty.
Einstein believed,
but could not prove, that although we didn’t know of a way to measure a
particle’s position and velocity simultaneously with great accuracy, the
particle nevertheless has a well-defined position and velocity, and it interacts
with other particles as if this is so. His speculation was described as needing
a “basement level” of physical laws, which had not yet been discovered. With
a “basement level” of physical laws the apparent “unknowableness” of a particle’s
properties would be just that, apparent “unknowableness.”
The particle “knows” where it is located and how fast it is going, and in
what direction relative to the rest of the universe - even if humans can’t
know.
This “quantum
weirdness” is often cited to discredit the idea that events are “determined.”
But we cannot rule out the possibility that future physicists will discover
a basement level of physical law, and that this will restore Newtonian physics
as a complete theory for all size scales. The new Newtonian physics would
have the old Newtonian physics as a first approximation, valid for use with
the vast majority of physical phenomena dealt with on a daily basis.
Starting here I
will present only brief summaries of chapter sub-sections that have been
moved to Appendix A for this Second Edition of Genetic Enslavement.
Levels of
Physical Explanation
The matter
of “levels of physical explanation” must be dealt with for the reader who
is not prepared to accept the existence of a basement level of physical law.
In the physical
sciences it is common to treat a physical process at a “higher level” than
atoms interacting in accordance with the most basic level of physical law,
a = F/m and quantum physics. Instead, other
“laws” are constructed for everyday settings, either derived from the basic
level of laws or derived from experiment and deemed compatible with the basic
laws. One example should serve to illustrate this.
Consider the
atmosphere, which consists of an immense number of molecules. Any thought
of using a = F/m applied at the level of
molecules for the purpose of predicting the weather would be silly because
of its impracticality. There is no way to know the position and velocity
of all the molecules in the atmosphere at a given time for establishing the
"initial conditions" required for subsequent calculation using a = F/m. The meteorologist employs a “higher
level of physical explanation” by inventing “laws” that govern such aggregate
properties as "atmospheric pressure," “temperature,” and "wind speed."
In each case
the invented property and rules for using it can be derived from a = F/m, so these handy properties and rule
for usage are “emergent properties” of the basic level of physical laws.
Every atmospheric scientist would acknowledge that whenever a meteorologist
relies on a handy rule, such as “wind speed is proportional to pressure gradient,”
what is really occurring in the atmosphere is the unfolding of an immense
system of particles obeying a = F/m.
Just because
scientists find it useful to employ "emergent properties" does not mean
that the emergent properties exist; rather, they are no more than a useful
tool for dealing with a complex system. A "pressure gradient" doesn't exist
in nature; it exists only in the minds of humans. Model idealizations of
an atmosphere can be used to prove, using a = F/m, that the thing called a "pressure gradient" is associated
with wind. But these very proofs belie the existence of the concept, for they
"invent" the concept of a pressure gradient for use in a model that then
uses a = F/m. The handy meteorology rules,
and their "emergent property" tools, are fundamentally redundant to a = F/m.
The refinements
of modern physics do not detract from the central concept of materialism,
which is that everyday (large-scale) phenomena are the result of the mindless
interaction of a myriad of tiny particles in accordance with invariant
laws of physics. Reductionists acknowledge the importance of the many levels
for explaining complex phenomena, but they insist that all levels higher
than the basic level of physical explanation are fundamentally “unreal” and
superfluous, even though the higher level of explanation may be more “useful”
than a lower level of explanation.
Science embraces
what might be termed the "first law of reductionism," that whenever a phenomenon
can be explained by recourse to a more basic level of physical law, the “higher
level” explanation should only be used when it is drastically simpler to
use and unlikely to be misleading. Whenever a higher level of explanation
is used, there should be an acknowledgement that it is being used for convenience
only.
Living Systems
Reductionists
view living systems as subject to the same physical laws as non-living systems.
Therefore, the behavior of a living system is an emergent property of a
complex physical entity. A living thing is thus an automaton, or robot.
The thing we
call "mind" is an "emergent property" of an automaton’s brain. The brain consists
of electrons and protons, and these atomic particles obey the same physical
laws as inanimate electrons and protons.
Such things
as "thoughts, emotions and intentions" are mental constructions of the brain
that in everyday situations are more "useful" than the laws of physics for
the study of behavior. In spite of their usefulness, they are not actually
causing the movement of particles in the living organism, and they don't
exist at the most fundamental level of understanding.
Even “free will” must be shorn of its essential features, and recast as another
"emergent" product of real causes.
“Consciousness,”
like “free will,” is also an emergent property of automatons, just as the
"wind" is an emergent phenomenon of the atmosphere. I don’t object to the
use of “consciousness” for the same reason that I don’t object to the use
of “wind” when an atmospheric science problem is to be solved.
It has been argued
that the physicist exhibits "faith" in extending what is observably true
in simple settings to more complicated ones. This assertion of faith is true,
but the faith follows from the physicist's desire to invoke a minimum of
assumptions for any explanation.
Some Practical Considerations
Concerning Levels of Explanation
The brain evolved,
like every other organ, to enhance survival of the genes that encode for
its assembly. It should be no surprise, therefore, to find that it is an imperfect
instrument for comprehending reality. If it is more efficient to construct
brain circuits for dealing with the world using concepts such as spirits
and prayer, rather than reductionist physics, then the "forces of evolution"
can be expected to select genes that construct brain circuits that employ
these pragmatic but false concepts. Since no tasks pertaining to survival
requires the a = F/m
way of thinking, the brain will find this to be a difficult concept. It is
a triumph of physics to have discovered that a = F/m and quantum physics rule everything!
A naive person might
believe that the primitive person, viewing everything in terms of spirits,
is thinking at a higher level than the scientist. This would be a ludicrous
belief. A primitive is a lazy and unsophisticated thinker. He is totally
oblivious to reductionist "levels of thought." As I will describe later, he
uses a brain part that is incapable of thinking rationally: the right prefrontal
cortex. Human evolution's latest, and possibly most magnificent achievement,
is the left prefrontal cortex, which evolution uses to usurp
functions from the right prefrontal cortex when rational
thought is more appropriate (i.e., feasible). Too often contemporary
intellectuals will unthinkingly succumb to the pull of primitive thought,
as when someone proudly proclaims that they are “into metaphysics" (an oxymoron).
A fuller exposition
of this topic cannot be given without a background of material that will
be presented in later chapters. For now, I will merely state that mysticism
is a natural way of thought for primitive humans. It is "easier" for them
to invoke a "wind spirit" explanation than the reductionist ones, such as
a = F/m, or higher level derivative physical
concepts. They do this without realizing how many ad hoc
assumptions they are creating, which in turn require explanations, and this
matter is never acknowledged (as with invoking God as an explanation, without explaining "God"). Their thinking
may seem acceptable from the standpoint of a right prefrontal cortex (or
"efficient" from the perspective of the genes that merely want to create
a brain that facilitates the gene's "goal" of existing in the future), but
it is terribly misguided from the standpoint of the thinker endowed with
a functioning left prefrontal cortex, that demands rational explanations
with a minimum of assumptions. This unthinking proliferation of ad hoc assumptions bothers the reductionist, but it doesn't
bother the unsophisticated primitive.
Reductionism is
for the Few
H. G. Wells must
have understood the issues of this chapter. The reductionist paradigm was
an important part of intellectual thought during the 19th Century, and Wells
grasped it more surely than even many scientists today. Scientists, engineers
and inventors must have been held in high esteem during the second half of
the 19th Century, and the first half of the 20th. The per capita number of
significant discoveries and innovations, as measured by Asimov's Chronology
of Science and Discovery (Asimov, informally distributed in the 1980s, formally
published 1994) peaked at about the middle of this period (actually, 1910
AD, as described in Chapter 15, and specifically Fig. 15.12).
Late in the 19th
Century, after Darwin’s evolution by
natural selection instead of divine guidance had time to register with intellectuals,
the idea of “humans as automatons” was part of the climate of opinion.
Thomas Huxley was intrigued by this idea, and Darwin humored him by
signing letters with a reference to it (Sagan and Druyan, 1992, pg. 70).
Reductionism requires that all living things be viewed as automatons, or robots
created by evolution. Yet none of today’s academics seem brave enough to
defend this idea.
Ernst Mach (1893)
deserves mention as an early champion of the idea that all branches of science
will eventually be viewed as unified. He was a continuing inspiration for
those who attempted to advance this perspective (Holton, 1993) throughout
the first half of the 20th Century. His was one of the most important
in a series of “flame-bearers” for keeping alive an idea that came out of
ancient Greece with the writings
of Democritus of Abdura (Sagan, 1980).
Reductionist ideas
were at least understood by literary people during the early 20th
Century. In 1931 novelist Theodore Dreiser, for example, wrote "I have pondered
and even demanded of cosmic energy to know Why.
But now I am told by the physicist as well as the biologist that there can
be no Why but only a How, since to know How disposes finally of any possible
Why." (Dreiser, 1931).
Sadly, we cannot
expect today's intellectuals to have the same profound understanding of the
nature of reality as was exhibited a couple generations ago by such writers
and social commentators as Wells and Dreiser. The quality of thought over
time, in a specific subject area, is not always progressive. As with civilizations,
there is a rise and fall in the sophistication of world views. Indeed, as
the 21st Century begins we are in the midst of a renewed interest in returning
to the comforts of primitive outlooks, as described in the next chapter.
─────────────────────────────────
CHAPTER
2
─────────────────────────────────
RESISTING THE BACKWARD
PULL
TOWARD OUR SPIRITUAL
HERITAGE
"O miserable minds of men! O blind hearts! In what darkness
of life, in what great dangers ye spend this little span of years! ... Life
is one long struggle in the dark." Lucretius, On the Nature of Things,
ca 60 BC.
"It does no harm to the mystery to
know a little about it. For far more marvelous is the truth than any artists
of the past imagined! Why do the poets of the present not speak of it? What
men are poets who can speak of Jupiter if he were like a man, but if he is
an immense spinning sphere of methane and ammonia must be silent?" Richard Feynman,
Lectures in Physics, Vol. 1, Addison Wesley, 1963.
The term "New Age"
is a misnomer, and an insult to better ages. It is a misnomer because it
is a regression to primitive ways of thinking, ways which should have remained
buried, yet which have resurfaced due to a mysterious mental pull toward
the primitive. This pull is unfortunately endemic to the flawed human mind.
"New Age" embraces the occult, a belief in angels, spirits, astrology, magic
and life after death. It is a return to the kinds of enslaving thoughts which
Lucretius urged his disciples to be rid of 2000 years ago.
The Primitive's
Reliance on Spirits
The environment
of our primitive ancestors, including both the physical and social aspects,
rewarded genes that constructed brains that could deal with the world, which
is profoundly different from stating that their environment rewarded brains
that could understand the world. As I argue in a later chapter, primitive
people did not employ the full powers of a modern left prefrontal cortex,
but instead relied upon a more primitive right cerebral cortex design for
both cerebral hemispheres. To the extent that "producing grandchildren" (a
convenient measure of genetic success) became more dependent upon mastery
of a world of human relationships instead of mastery of the natural world,
the architecture of the human brain evolved in ways that favored comprehending
the social world at the expense of the natural one.
The social arena
is less predictable than the natural one, so different mental abilities were
rewarded in an environment requiring social skills. When a brain that evolved
for the social setting addresses matters in the inanimate world, it should
not surprise us to find that such a brain employs "weird logic" in this
neglected realm. The primitive's vision of the world, being unguided by
rational thought, was filled with spirits that behaved like people. Primitive
people have gods for lightning, wind, rain, light, dark, and whatever seems
important to a primitive's precarious life. Thus, when the sun god loses
a conflict, according to this weird logic, it follows that there shall be
wind, rain and lightning.
Today’s common belief
systems provide evidence that for our ancestors the need to competently deal
with human affairs was more important to the evolving human genome than the
corresponding need to competently deal with the inanimate world. In high‑tech
modern Japan, for example, the
indigenous Shinto cult and religion remains popular. Shinto worship centers
on "a vast pantheon of spirits, or kami, mainly divinities personifying aspects
of the natural world, such as the sky, the earth, heavenly bodies, and storms.
Rites include prayers of thanksgiving; offerings of valuables..." (Encarta Encyclopedia, 2000). Even well‑educated Chinese
still believe in Feng Shui (the need to please spirits by a proper placement
of furniture, entrances, etc). American Indians, who crossed the Bering Strait 13,000 years ago,
brought with them a burdensome need for believing in spirits that demanded
ritual obedience. There seems to be an abundance of depressing examples from
every culture.
The Dyads of Primitive
Thinking
Dyads abound in
primitive thinking. Night and day, good and bad, friend and foe, birth and
death ‑ they all contribute to a "yin and yang world." It is not surprising
that when primitive men floundered to explain the world, they relied upon
a dyadic competition. Thus, night and day are engaged in a daily struggle,
literally; and at sunrise the "day" has become victorious over "night," and
so on. But it gets complicated, for during winter the stronger competitor
is night, whose exhaustion gives day the upper hand during summer.
Conflict permeates
a primitive's thinking, because conflicts between tribes define primitive
life. Nevertheless, men battle upon a stage set by even stronger forces than
themselves. The weather is overwhelmingly strong, as is the ocean, the occasional
earthquake, tsunami and volcano. There must be gods in heaven who unleash
the thunderstorm and lightning, that punishes
and rewards men. Since powerful men can be appeased, or slightly influenced,
so might the gods. Man's quest for control over his fate led him in false
directions, for gods cannot be appeased when they don't exist.
We should laud the
primitive's urge to explain, even though it seems to be only weakly motivated
by an urge to understand. The human claim for nobility rests upon this urge.
But let us also not be mistaken about the explanations created by primitive
men: Primitives have been stupendously wrong in almost every instance!
Their explanations
were wrong because they arose from a primitive right brain. Only recently,
with the ascendance of the aforementioned, fast‑evolving left brain, with
its logical mode of thought and lack of traditional "wisdom," has it been
possible to conjure up correct explanations. But, so strong is the irrepressible
right brain that even many contemporary "intellectuals" still believe that
primitive explanations contain some profound and subtle wisdom that makes
it "just as valid."
Thinking men of
every age seem to have sensed a pull toward primitive thinking, and worse,
toward primitive behaving. The decay of civilization has been an ever‑present
concern for those who live in a civilized state. This concern was expressed
by ancient Greek philosophers, just as it is in today's world.
We know that the
civilized state is not secure, because we sense the presence of that insistent
and primitive right brain. To use the primitive's own metaphor, we are engaged
in a struggle between good and bad, between light and dark, and it is now
"late afternoon." Some of us who worry about the
approach of evening, and a long night, admonish our contemporaries to resist
the "primitive pull," to stay the course that brought us to this glorious
noon, atop the highest
mountain, by keeping the new faith as it struggles with the old. It has
become a battle between the two titans of human history:
the two brain halves!
The Modern's "Spiritual
Cleansing"
The primitive way
of thinking is more efficient to implement than the modern physicist's cumbersome
a = F/m and quantum physics way of thinking.
We moderns smile
at those faltering attempts to see order in nature. From our 21st
Century perspective, we see that their "explanations" are pathetically simple‑minded,
and emphatically wrong!
Yet most people
today feel comfort in being pulled in this primitive direction. It's as if
the more complicated and up‑to‑date explanations require too much effort,
resisting as they must the objections of old brain circuits. The result,
for most people, is that the brain maintains old and new understandings side‑by‑side.
The human brain is amazingly adept at compartmentalizing thought, and allowing
the most irrational beliefs to coexist beside enlightened ones. "Cognitive
dissonance" is minimized by insulating brain circuits from each other.
More than a few
scientists surrender rationality on Sunday. I once worked with a scientist,
a master of magnetic fields on planets in our solar system, who believed
in the many levels of heaven taught by the Mormon Church. I give more examples
of this in Chapter 13.
During Humanity's
long march to the present, we have progressed from "magic" explanations to
"rational" ones. Those brave thinkers who led the march have shed the magic
and embraced the rational. Rationality led to reductionism, which I believe
is Humanity's greatest intellectual achievement! The march forward has been
led by people whose style of thinking adheres to the values of our left
cerebral hemisphere, or left brain. The regressive, backwards pull is from
a majority of "neurologically primitive people" whose thinking style remains
right‑brained.
Humanity's path
to reductionism has been "forced" by necessity. Imagine the consequences
of taking your car to a minister for its repair, instead of a car mechanic;
or seeking medical help from a shaman medicine man instead of a medical doctor
or nutritionist. Our primitive ancestors had no need for car mechanics, and
in their time medical doctors didn't exist, so they had less to lose by
adhering to magic. With the unfolding of time, and the accumulation of technology,
there has been a growing need to distinguish between spiritual and rational
explanations.
Not all aspects
of modern life require rational explanation. My friend who believed in many
levels of heaven was unencumbered by this belief during his work hours. His
spiritual beliefs might, in fact, have had a stabilizing effect in his personal
life. It is relatively inconsequential whether a person believes they will
go to heaven when they die, or dissolve to dust! It is more important that
they know about family budgets, cars, computers, and nutrition.
Whereas it may not
matter to a person's success at living whether all remnants of spirituality
have been purged from his intellectual outlook, it does matter to the person
engaged in a serious endeavor to understand "the nature of reality." Every
serious thinker is obligated to undertake a lifelong vow to cleanse away
all vestiges of spirituality!
We must control
the impulse to regress to a world of spirits, no matter how comforting it
may be. As argued by Lucretius, we must move forward, abandon belief in personal
guardian angels and protecting gods, and replace them with understandings
based on rational thought.
Are We Making Progress?
During the past
80 years scientists have slowly aligned their personal beliefs with rationality.
In 1916 Leuba surveyed the beliefs of 1000 randomly selected scientists and
found that 42% of them believed in God, whereas a similar survey conducted
in 1996 showed only a modest decline to 39%. The belief
in immortality declined by a slightly greater amount, from 51% to 38%.
Perhaps more revealing, the more accomplished
scientist is less likely to believe. Leuba (1914) surveyed 400 "greater" scientists
and found belief in God to be 28%, whereas Leuba (1933) found that 19 years
later the belief rate declined to 15%.
Today, Larson and
Witham (1998) report that among 517 American scientists who belong to the
prestigious National Academy of Sciences only 7.0% believed in God. Considering
a belief in immortality, the above studies report that for 1916, 1933 and
1998 the belief rate was 35%, 18% and 7.9%. Among the general population
of non‑scientists, 96% believe in God!
There appears to
be a decoupling of what people of accomplishment believe and what the hoi poloi believe. Thus, among "greatest" and "accomplished"
scientists the rates of belief in God and immortality are low and declining
dramatically, among the ranks of scientists as a group the belief rates are
less than half and declining slowly, whereas among the general population
the belief in God is high and remains unchanged. Only the intellectuals are
abandoning God!
Faltering Progress
As my chapters on
the brain explain, I believe that the right prefrontal cerebral cortex finds
mysticism and religion congenial to its way of thinking, whereas the left
prefrontal cerebral cortex is inclined to think rationally. There is little
doubt that the left cerebral cortex is a more recently evolved brain area
than the right, as it is responsible for speech, conceptual thought and logical
thinking. The practice of science requires a well‑developed left brain, although
a well‑functioning right brain is also required in a supporting role. I speculate
that scientists typically have left brains that "dominate" their right, in
the sense that the left brains use the right brains as "tools" in the pursuit
of left‑brain‑directed activities. The scientist values things that the left
brain values, and the scientist's approach to studying a problem, and the
standards of proof, are consistent with the style of thought of a left brain.
Among the others, it is the right brain that employs its left as a tool,
keeping it subservient to right‑brain values and goals. This more common
style is a phase humanity must continue to "evolve
through" if it is ever to reach a winning place as a sentient species.
As I look back upon
the recorded history of the human groping for an understanding of who we
are I discern good and bad eras. The first good era was 5th Century
BC Greece, when the Ionian
philosophers articulated the reductionist paradigm, as described by Sagan
in his book Cosmos (1980, pg. 175). Democritus was the shining
star of that era. The next era would be 1st Century BC Rome, when Lucretius
wrote his famous poem On the Nature of Things. There followed
a Dark Ages millennium during which anyone who thought rationally had to
keep their thoughts secret. Stirrings of rationality started in 17th
Century Denmark. The 18th
Century Philosophes in France produced a full-bloom
resurgence of interest in Greek thought, but this rebirth was short-lived
due to the French Revolution. During the 19th Century European
discoveries buttressed support for rationalism and the reductionist paradigm
in particular. The new physics of the early 20th Century began
to discredit reductionism, unfairly in my opinion, though rationalism continued
to prevail. The global depression quieted many independent thinkers (like
H. L. Mencken), and this marked the beginnings of a slow return to spiritualism.
At this writing, in 2006, I see only a downslide of critical thinking that
cannot compete with the primitive appeal of religion and an excess of politically-correct
embrace of diversity of thought, no matter how irrational. A new Dark Ages
might be approaching.
Laying a Groundwork
of Understanding
The prospects are
poor that during the next century the general public will embrace a rational
outlook. Most people, even most intellectuals, are inclined to use the term
"reductionism" disparagingly. This can be understood if their belief systems
are influenced by a primitive, right prefrontal cortex. Rationality, which
is a left prefrontal cortex creation, is in conflict with the old right pre‑frontal
cortex. A fuller explanation of this will follow chapters on human evolution,
the brain's role in human evolution, the appearance of the artisan, his
role in the rise of civilizations, and the resentment of the artisan's rapid
rise to power.
These chapters,
in turn, will be preceded by a tutorial on genetics, using the sociobiological
paradigm. This will be our task for the next three "genetics tutorial" chapters.
If, dear reader,
you find the genetics tutorial chapters tedious, then skip them if you must.
You merely risk not having some tools for understanding the "micro‑motives"
underlying the "macro‑behaviors" under discussion. The genes, after all,
underlie everything pertaining to life!
The chapters describing
human evolution (Ch. 6) and the brain's role in human evolution (Ch 8) are
intended to illustrate reductionist ways of thinking about the evolution
of human nature, and should not be skipped. They provide a background for
understanding the following speculations on the rise and fall of civilizations.
The utopias and
living wisely chapters will resume the main theme of this book, which is
concerned with the individual's predicament of living with "outlaw genes."
The intervening chapters present a story of how humans came by the weird human
nature we're stuck with, and I see no way of resuming the main topics without
the preparation of these intervening chapters.
─────────────────────────────────
CHAPTER
3
─────────────────────────────────
GENETICS TUTORIAL
‑ PART I
"...organisms die
but their genes pass on ‑ often mutated and redistributed, it is true,
but genes nevertheless; and it is difficult, therefore, to escape the conclusion
that the design of the organism is merely to provide for gene multiplication
and survival..."
Carl Sagan, "Radiation and the Origin of the Gene," Evolution,
January, 1957.
People once believed
that the universe was created for Mankind and all other life was placed here
for our use. This was gradually replaced by the harsher belief that our
species competed with other species, and that Human Nature was designed to
do what was good for the species. Anyone who acted selfishly was aberrant,
and would be punished in a later life. Darwin believed that
individuals competed with each other, and the victors of individual conflict
shaped human nature. But now, the level at which competition occurs has descended
one more level: sociobiologists argue that gene alleles compete with each
other for positions on chromosomes (W. D. Hamilton, 1964a,b; G. C. Williams, 1964; E.O. Wilson, 1975; R. Dawkins,
1976).
If the combatants
are the genes, then what are we individuals? We are the "lumbering machines"
carrying the genes that assembled us for the genetic competition (Dawkins,
1976). An individual is like a puppet, whose behavior is directed by strings
that are pulled, ultimately, by tiny genes (please excuse the poetic license
and anthropomorphism of this phrasing). The demotions our egos suffered during
the past couple centuries continues into the 21st, as people
must now deal with the thought that we are created by our genes for gene
battles, and the genes do not care about the individual's welfare.
If the genes are
this important then we should know their story, from the beginnings of life
to the present. I shall present a recapitulation of the evolution of life
on earth, with a proper emphasis on the role of genes. Some of these descriptions
are speculative, yet illustrate ways that I believe the subject should be
approached. The essence of every speculation is, of course, mechanistic reductionism!
A Brief History
of Life
When earthly life
started, 3.5 billion years ago, tiny replicating molecules resembling DNA
(or maybe RNA) must have competed with each other for incorporating their
molecular building blocks into copies of the replicating molecules. In time
those molecules that accidentally created a protective "coating" survived
longer. This crucial event might have been hastened by the existence of water
droplets that would naturally form surface layers of like‑charged molecules
with hygrophobic ends (Donaldson et al, 2004). A droplet
with such a covering is a rudimentary cell, as the cell "wall" may have protective
properties.
Approximately 2
billion years ago, a well‑functioning one‑celled form appeared which housed
cell‑creating DNA floating inside (prokaryote). Later, a variant of the prokaryotic
one‑celled life form had the several DNA molecules confined to a cell nucleus
(eukaryote). This represented one more structural level of protection of
the DNA. Whereas it must have helped the DNA survive it also required that
a solution be found for the slightly more complicated replication process.
A trend is evident,
and it has continued throughout the long story of life on Earth. To compete
better, DNA molecules have had to wrap themselves in an ever‑more complex
structure, devise ever‑more complex methods for replication, and retain control
of their protective structures for competition with other life forms.
Multi‑celled creatures
did not appear until about 1 billion years ago. Each cell contained a nucleus
with an identical set of genes inside. It is possible that initially all
cells in a multi‑celled life form were identical. A grouping of cells has
a smaller "surface area per mass" exposed to the watery environment than one‑celled
forms, and this added protection may have rewarded the forms that tended
to stick together.
Specialization of
some cells in a many‑celled creature may have been the next step in the evolution
of life. With a reliable association of cells having the same DNA, there
existed a reward for any gene mutations that helped the outermost layer of
cells to specialize in protective matters. Since all cells had the same nucleus,
it could still be argued that the sacrifice of a few cells to become mere
"protective skin" while forsaking reproduction themselves nevertheless enhanced
the reproductive prospects of the identical genes inside the cells that were
being protected. This concept is a kernel for "inclusive fitness," described
below.
In this way, skin
might have been the first "organ" to evolve. Once a method had evolved for
guiding a cell's properties to be responsive to its surroundings, the path
lay open for the evolution of any number of organs. Organisms competed with
differently constructed organisms in seeking food - and perhaps in consuming
each other. Although the organisms competed with each other for food, and
perhaps attempted to destroy or devour each other, since the fate of this
competition was determined by the properties produced by the genes within,
it is more insightful to view the competition as occurring between the various
gene groups than between individuals.
Gene Competition
Within and Between Species
Specifically, only
the genes that differed were in competition. Identical genes in competing
organisms might appear to be in competition, but only because they were part
of an organism that had different genes. Since the fate of identical genes
in different organisms was not in question, they were not competing with
each other. If organisms had chromosomes that differed at only one gene location,
and only two gene forms (alleles) existed at this location throughout a
population of organisms, then it would be the two genes (gene alleles) that
were in "competition."
The casual observer
who thought that two kinds of organisms existed that were competing with
each other (for food, let's say) would be misunderstanding
the situation. A deeper understanding shows that two genes were competing
with each other.
The word "competing"
is a human‑invented concept. It is important to remember that the entire
process of genes competing, with one gene mutation slowly yielding to another,
is purely mechanistic. Obviously, the genes are unaware that they are "competing."
Only a human observer would remark "these two genes are in competition."
Individuals within
a species may "compete" with each other, and so may individuals belonging
to different species. The underlying dynamic is the same:
every gene acts as if it wants to proliferate and last forever. Again,
a gene does not "want" to proliferate. Rather, those that in fact proliferate
are the ones that express themselves as individual characteristics that the
human mind will identify as "wanting" to proliferate. Thanks to our primitive
right brain (that evolved for dealing with social settings) it is helpful
to use social metaphors for explaining mechanical processes.
It is theoretically
possible that two species could exist in competition with each other while
having no genes in dispute within each species. In other words, all individuals
of one species would be genetically identical, and the same for the other
species. Yet, since we are supposing that they are competing for the same
resources in their environment, for example, the two species are in competition
with each other. We should further specify that the two species hold some
of the same genes (this is very likely, since they have common ancestors).
For this hypothetical situation, two large chunks of genes are in competition
with each other and the "winner will take all" after one species is exterminated
(which is common for species that occupy identical niches).
This is one extreme
of a continuum of situations. At the other end is a situation in which there
is no between‑species competition, but there is competition between genes
within the single species. In other words, some gene loci on the chromosomes
have two or more alleles, and these alleles are in competition with each
other. The simplest case would be one gene locus with two alleles, and the
two alleles are competing with each other for exclusive presence at one locus
on the chromosome.
This simple situation
is likely to end with a complete win by one of the alleles. However, there are special cases where there will be a
steady‑state percentage representation of both gene alleles (i.e., an evolutionarily stable strategy, or ESS, as described
originally by J. M. Smith and later by Dawkins, 1976).
In the real world
there will be some within‑species competition of counterpart gene alleles
and some between‑species competition between all the genes that are different
between the two species. For the between‑species competition, large chunks
of genes are involved; but only some of these will be under the influence
of selection pressures.
Thus, for the typical
situation, some gene alleles will be changing their representation frequency
within the species gene pool due to "other species" selection forces, while
other gene alleles will be changing their representation frequency within
the species gene pool due to "within species" selection forces.
Appendix B presents
examples of the ruthless nature of gene competition using human virus examples.
Inclusive Fitness
The individual organisms
called humans should be forgiven for seeing everything in terms of what they're
good for to the individual organism. Dawkins (1982) describes an aberrant
period in biology when the paradigm shifted from a "selfish organism" perspective
(starting with Darwin) to "adaptation
for the benefit of the species" paradigm (during the mid‑20th Century).
Given that many (unsophisticated) people believe that people do things that
benefit the species, the level at which people perceive evolutionary competition
to occur should shift in the other direction, leading eventually to the "selfish
gene" paradigm.
In other words,
if we seek insight into how evolution works we should not ask "what good
are genes to individuals?" but "what good are individuals to genes?"
An example will
serve to show how ridiculous an imperfect paradigm can be. A graduate student,
who must still retain remnants of the belief that adaptation is for the good
of the individual, studied the Australian redback spider. She describes how
the male positions himself during copulation so that the female can eat
his body during a leisurely insemination process, thus satisfying her into
not seeking another male. This assures that the sacrificing male's sperm
will not be competing with the sperm of another male. To sum up the article,
the magazine author presents the following astounding quote attributed to
the student: "Sexual cannibalism has always been thought of as a conflict
between males and females, in which males are just being overpowered. It's
important to realize that it can be advantageous for the male."
Only someone handicapped
by the "good of the organism" paradigm could say such a ridiculous thing!
The benefactor, clearly, is the gene that causes the male to behave in this
bizarre manner, not the individual male. He is a victim, more than the female.
What's missing in this summarizing statement is an acknowledgement that genes
produce behaviors which can manipulate and victimize individuals with behaviors
that benefit only the genes that create the behavior?
Inclusive fitness
states that genes tend to produce individual behavior which maximizes the
presence of the behavior‑producing gene in succeeding generations (Hamilton,
1964a,b). Since biological relatives are likely
to be carriers of the same genes as the behaving individual, the fate of
the genes in relatives matters as much as the genes in self. For social animals
that live among relatives the merit of an action must take into account the
consequences upon those "genetically related" carriers of the genes.
The mathematical
treatment of inclusive fitness is straightforwardly presented in many places.
I shall simply give a feeling for it by repeating a traditional example.
A gene will reward a self‑sacrificial act if it saves at least two cousins,
or at least four second cousins, etc., as in each case the
same number of identical genes is preserved (on average).
Inclusive fitness
provides an explanation for "altruism" by the argument that an altruistic
act promotes the proliferation of genes generating the act. Sometimes these
altruistic acts are at the expense of the individual; but that's OK from
a gene's perspective, for the individual is merely a tool created by the genes
for gene proliferation!
The inclusive fitness
paradigm is useful in many other situations, some of which will be dealt
with in the following chapters.
─────────────────────────────────
CHAPTER
4
─────────────────────────────────
GENETICS TUTORIAL
‑ PART II
"Thus the earliest
vertebrates, like the earliest amphibia, the earliest mammals, and the
earliest primates, were small predators. Over and over again in evolution,
the originators of new modes of life were small predators, and the key innovations
at each stage conferred a selective advantage in predation." John Morgan Allman,
Evolving Brains, 1999, p. 73.
In this chapter
I continue describing some basic principles of evolution that apply to all
living things. It will serve as a foundation for the more speculative and
interesting evolutionary results found in human nature.
Pre‑Adaptation
Some genes are "pre‑adapted"
for new environments. A gene is pre‑adapted if there was a negligible reward
for its presence in the genome at the time some new environmental challenge
appears for the first time, and for which the gene then confers a significant
genetic benefit.
Modern society provides
many examples. Computers didn't exist before the mid-20th
Century, yet we find that many individuals
are naturally talented for computer programming, design, networks, and other
aspects of computer use. These people have genes that are pre‑adapted for
the computer environment.
Pre‑adaptations
are always present, as the following thought experiment illustrates. Imagine
any task, and a procedure for reliably measuring performance of that task.
The task could be jumping as high as possible, or remembering a sequence
of numbers ‑ any task will do provided performance can be measured objectively,
producing a continuous range of scores (a binary result, such as pass/fail,
does not meet this "continuous range of scores" criterion). After two people
have performed the task, there will invariably be a "better" and "worse"
performance. After many people have performed the task, the test scores may
form something resembling the Gaussian, or "bell curve" distribution, with
many scoring near a middle region, and fewer scoring really well and poorly.
The top scorers can be described as "pre‑adapted" for the task (provided the
task is novel or evolutionarily "new").
In real‑world situations,
whatever the change in environment, whatever the change in job opportunities,
whatever new sporting games are invented, there will always be new consistent
winners and losers. Winners in the new task might have been mediocre performers
in the old ones (and old winners may become the new losers).
"We are what we're
good at," and the forces of selection measure us by what we're good at in
the context of our times. Whereas the computer whiz is pre‑adapted to this
era, so might a “nobody” of today be pre‑adapted to some future era. We should
be careful in judging others, for they might have shined outstandingly in
past settings, or be examples of a type that will shine in future ones. Faceless
nobodies of times past might have rivaled the best of today's stars, if
only given the chance by a change of environment. Chance is everything!
Pre‑Maladaptation
But if the winners
are pre‑adapted, the losers are "pre‑maladapted." It may seem "unfair" to
a civilized mentality to believe that "pre‑ordained" winners and losers
will exist when new opportunities appear, but every species has been molded
by this unfair rewarding of individuals through abrupt environmental change.
We may not like it, but this is the way things work.
Anyone feeling gratitude
for evolutionary accomplishments should also feel thankful for the diversity
of individual performance. Thanks to "inequality" evolution proceeds! But
while we celebrate inequality, and rewards for the pre‑adapted, let us also
have compassion for the pre‑maladapted, the world's ill‑fated losers, for
they cannot be responsible for the changes that doom them.
Over and over, in
this book, we shall encounter repugnant examples in nature. Our lesson is
to accept that Nature doesn’t care about individuals, only the genes! And
the genes have no qualms about wasting individuals for their sake. Fish lay
thousands and millions of eggs, so that on average one or two will survive.
Several insect species produce male brains that are programmed to allow the
female to make a nutritious meal of him after copulation ‑ to postpone her
mating with a competitor or for nourishing his offspring! The historical
record shows that humans will send legions of young men to battle, like fodder,
who in the prime of their life become maimed or killed. The victors in battle
rape the vanquished men's women, then march home
as heroes, with greater rights for domestic breeding. In all these settings,
the individual is "sacrificed," for he is engaged in risky behaviors with
benefits that accrue reliably to only the genes.
Humans who ponder
the consequences of what I'm calling a pre‑maladaptation have grounds for
bemoaning their bad luck. I like the thought that each person "has their
time," a time when they would have some maximum of pre‑adaptation, and since
people are born into times "at random," they most often are "out of their
preferred time." Imagine how frustrating it would have been for Beethoven
to have been born before pianos existed, and before orchestras. Or for Einstein
to have lived before the preliminaries of 19th Century physical
theory had been set. Delay Darwin's birth a century;
would he have become the giant we know today? Bring to the 21st
Century such notables as H. G. Wells, Lucretius, Democritus, Shakespeare,
Homer, and others; what would become of them? We cannot know how fortuitously
attuned to their age these giants were, or what nobodies they might have
been had the "roll of their genes" occurred at some other time.
Species Shaping
Forces
Pre‑adaptation is
a useful concept calling attention to the fact that whatever an organism's
make‑up it will have some kind of “match” to every hypothetical new environment,
and the match of some individuals will be adaptive. It might be adaptive
whether or not the environment in question has ever existed before, and whether
or not any of the organism's ancestors have been exposed to that environment.
In such cases, we should not say that the organism has become adapted to
the environment in question, just because it fares better than some of its
cohorts. Rather, it is adapted due to a pre‑adaptation.
Most organisms will
be pre‑maladapted to the new environment. Thus, most individuals will
fall behind, watching a minority of pre‑adapted individuals leap forward.
The greater the number of changes to the environment, the greater will be
the disparity in relative rewards between the pre‑adapted and pre‑maladapted.
A species should evolve "faster" at such times.
When I refer to
changes in the "environment" I mean to include not only the climate for a
region, and the disappearance of a food staple (plant or animal), or appearance
of new foods, but also the appearance of a new predator, the invasion of
a new parasite, or the adoption of a new element of "culture" (in the case
of humans, and perhaps chimpanzees).
There is a special
case, unique to humans, in which culture has created
an entirely new environmental condition: the removal
of most of the natural threats to survival.
An advanced civilization
shields people from diseases, animal predators, and, in some cases, the need
to work. It even shields people from each other to a great extent, by reducing
the frequency of outbreaks of "tribal warfare." In this environment genes
that in harsher, unforgiving environments would be maladaptive would
now be neutral. Only the most severe genetic defect will be eliminated from
a human genome shielded this way.
Under these conditions
we might want to think in terms of "potential pre‑adaptation" and "potential
pre-maladaptation." Today's genome is accumulating a large reservoir
of potential pre‑maladapted genes, carried unknowingly by individuals who
may be reproductively successful only because they are not subjected to selective
forces.
At the risk of getting
ahead of my story, I believe that such genes will become apparent only after
natural forces of evolution are restored, and put "the squeeze" on our burgeoning
global population. Winners and losers in this new environment will not be
close‑call winners and losers, they'll be clear‑cut
winners and losers. The disparity between those now destined to win and those
destined to lose is greater than ever, and growing
faster than ever. The complexion of Humanity could change dramatically apre le deluge.
To understand a
species we must consider the selective forces that have "shaped" it. In other
words, we must learn what kills individuals before they reach reproductive
age, what factors determine which individuals reproduce after reaching maturity,
what foods are eaten, and how precarious is the supply.
For example, if
our ancestors 5 million years ago were eaten by lions the survivors would
have been good at avoiding lions. This might have rewarded the evolution
of bipedality, which would have enabled standing tall and running fast. It
might also have rewarded the capacity for social cooperative strategies, a
precursor to intelligence.
Another theory speculates
that our ancestors had to learn how to find and store root plants that would
have grown on the grasslands (Wrangham and Peterson, 1996). This would have
rewarded the creation of digging tools, and the ability to carry extra roots
to a storage place at a home base, which in turn would have rewarded bipedalism
and a self‑control that provisioning requires.
Whichever environment
accompanied the branching of bipedal chimpanzees from their jungle‑dwelling
forebears 5 million years ago, we can be sure that the forces of selection
rewarded individuals carrying genes for dealing with whatever were the causes
of mortality in their new environment, whether they were escaping from lions
or digging and storing roots.
Perhaps 500,000
years ago some humans migrated to the edge of constantly‑moving glaciers.
Mortality in this new setting would have been climate‑related, such as cold
and hunger. We may presume that genes for planning and foresight were rewarded.
To the extent that large animals were hunted, and meat became an essential
food source, genes for a strategic type of cooperative hunting would also
have been rewarded.
After the last glacial
cold period, that peaked 19,000 years ago, humans had to adapt to an ever‑warming
climate. For some, this meant adopting an agricultural lifestyle. Those who
were pre‑adapted for farming would have prospered, provided they could also
band together for mutual protection from raiders. Others remained nomadic,
and targeted the new farmers. Thus, the main killer of Man became other men
(it probably has been "other men" for the past 100,000 years, at least).
As farming achieved unprecedented success, urban living became possible, sometime
after 3000 BC. This created opportunities for microbes, which competed with
Man as the main killer of men. Our ancestors are the ones with immune systems
that afforded protection against "urban" diseases.
In every step of
this evolution toward modern Man, the change in what killed people was a
principal selective "force."
H. G. Wells made
the point, 100 years ago, that long‑lived life forms cannot adapt to fast
changes of conditions, unlike short‑lived forms, that can adapt. This leads
more often to the demise and replacement of long‑lived large creatures by
other large creatures, both of whom are competing with small, short‑lived
creatures. He warns that humans, with a long life span, are vulnerable on
this account.
The looming threat
to Humanity posed by viruses and bacteria may become a classic example of
this evolutionary dynamic. How ironic if our demise, or loss of greatness,
which is most often portrayed in terms of dramatic events, such as global
thermonuclear war, instead is dealt by tiny viruses!
If in fact viruses
produce large‑scale human die‑offs during the 21st century (Garrett,
1995), the survivors will be those with pre‑adapted immune systems; not the
physically strongest or most intelligent. This possibility illustrates in
dramatic fashion the principle that "a species is shaped by what kills its
members."
How Many Genes Can
Compete?
Human tribes are
supposed to have numbered 50 to 100 individuals throughout much of our prehistory.
The number of adults in such a tribe would have been about half this number,
half of whom would have been adult males (12 to 25 in number). It is tempting
to think that fewer than this number of genes can be evolving in the tribal
genome. But such an assumption is erroneous, as I will illustrate.
When a man goes
into the world "to be measured," it is his phenotype that
is being measured. And his phenotype could be the result of many genes (interacting
with each other to produce a unique phenotype). Consider the extreme case
where each of the men in a tribe differs from "an average" by just one allele.
Consider 4‑year intervals, during which each woman of child‑bearing age bears
one baby. During each 4‑year interval, if only one man prospers and is accorded
sole breeding status, then every 4 years one allele can be declared a winner
over it's competitor(s). In a lifetime, 10 alleles
can be declared winners (where we imagine the environment places great importance
upon different aspects of phenotype each 4‑year period). After 80 years,
20 alleles could be declared winners, etc.
Even though this
is a thought experiment, it proves that there is no fundamental, mathematical
reason forbidding the number of gene sites for allelic competition to exceed
the number of adult males in a self‑sufficient tribe, or the total number
of tribe members. (A "multiple regression" statistical
argument is also possible, and more persuasive for me, but I shall spare
my readers of this daunting argument.)
Every individual
is a carrier for many genes that are competing with allele counterparts.
The number could be 50, or 500; and it doesn't matter if the individual is
a member of a tribe with only 50 or 100 members.
Migration, New Gene
Competition, and Pace of Evolution
The number of gene
loci hosting allelic competitions has undoubtedly increased in number since
the advent of urbanization, and the more recent globalization of our species.
A tribe of Africans may be homozygous for genes influencing skin color, but
if they were captured and brought to America as slaves their
ancestors would find that those same genes influencing skin color had become
a factor in determining individual welfare in the non‑African society. The
same argument would apply to many genes.
The inescapable
conclusion is that the more diverse a population becomes, due to migration,
the more genes there are in competition with each other. Does this mean human
evolution is progressing faster today, or slower? If a person's "measure"
is affected by more genes, it must take longer for all genes to have their
measure taken. Stated another way, when more genes enter the fray of competition,
those already in competition may feel a decrease of selective pressure influencing
their fate. Aspects of a person which were important in the tribal setting
suddenly recede in importance, as other genes, which had been firmly established
many generations earlier, resume their competition with alleles they had
never before encountered, or had encountered long ago and had triumphed over.
The coming together of ethnicities must introduce major changes to the set
of genes that are subject to evolutionary forces, in terms both of which genes
are in competition and the relative strength of selective forces upon specific
genes. These considerations suggest that the pace of evolution has slowed
in modern times.
Another slowing
influence is the declining rate of infant and childhood mortality. Unless something important has been overlooked, these arguments
suggest that the pace of evolution has slowed in modern times (Kondrashov,
1988).
Conservation of
Selective Pressures: Pleiotropy and Polygenes
When selective forces
suddenly reward a new capability the species undergoes a quick disintegration
in other, more recently‑acquired capabilities. This is due to the random,
unintended deleterious effects that any mutation produces, which places a
brake on the speed with which new capabilities can be acquired.
To understand this,
recall that a gene has many effects, referred to as pleiotropy. This is most dramatically illustrated when a mutation occurs
that has no redeeming consequences. For example, one mutation causes its
carrier to have 6 fingers, short stature and heart murmurs (Ellis‑van Creveld
syndrome). These phenotypic effects are seemingly unrelated, yet they are
caused by just one allele. Mutations that are adaptive, judged by the fact
that they have been selected during the course of evolution, will also have
many effects, with perhaps just one of them being adaptive to a far greater
extent than the numerous, small negative effects.
Thus, whenever a mutation occurs and confers an increment of adaptive advantage,
its future in the gene pool will depend not only on how well it performs
its adaptive task, but also upon how many unintended, deleterious effects
come with it.
Assuming for the
moment that there are only 40,000 genes in the Human genome, since there
are more than 40,000 properties defining a human, each gene must have more
than one beneficial effect. This implies that after a gene is "in place" it
can be modified over time to produce more desired effects. An "old gene" may
thus have several beneficial effects, in addition to a few small negative
ones. The selection of a modified, dual‑purpose (or multi‑purpose) gene must
occur with "painful slowness" since the original function of the gene should
not be disturbed appreciably, and since every mutation is likely to produce
other unwanted effects. To get things "just right" must require many generations
and many small compromises.
Whenever a new selective
force becomes important, the other selective forces must lose importance,
else the population will drop to dangerous levels. This "partitioning" of
selective pressure leads to a more conservative behavior of our genome, causing
already established gene alleles to remain longer than otherwise.
By the same reasoning,
a recently‑established gene allele is more likely to be disrupted with deleterious
effects than a long‑established gene allele. This "genetic entrenchment"
is due in part because of the rewards of redundancy for genes that are important
enough to respond to selective forces for long periods of time. A task that
must be done by a gene will be less vulnerable to mutation to that gene if
it has been exposed to mutations and selection for a long time. In addition,
genes that exist for a long time may become "depended upon" by other genes
that are selected after the first gene and which in some way depend upon
the presence of the first gene for its new effect to be expressed properly.
When two or more genes must be present to produce a specific phenotypic trait
that has adaptive value, those two or more genes are referred to as a "polygene"
group. Genes that are members of a polygene are more difficult to get rid
of, provided they have not become harmful. New genes have not had this opportunity
to achieve robustness, or become entrenched, and they are thus more likely
to be lost by random mutations because they are likely to have small phenotypic
consequences. The concept of "genetic entrenchment," and a culturgen counterpart
to this concept, is treated at greater length in Chapter 16.
Brain Genes
For humans it has
been estimated that at least 20% of the genome influences the brain. This
is not to say that 20% of human genes are present exclusively for brain wiring,
since many genes will exist mainly for other purposes which have "acquired"
brain wiring roles. If one of these genes mutates it is more likely to affect
its new brain task than the older, original task. Undoubtedly some genes
are mostly brain‑related, and probably some genes are exclusively brain‑related.
Whether a gene is partly, mostly, or exclusively brain‑related, if it recently
acquired this role it is likely to be more vulnerable to random mutation
than the other parts of the genome, or to older genes that mostly affect
anatomy or physiology.
Parts of the modern
human brain evolved during the past 100,000 to 200,000 years, and some people
speculate that for the past 40,000 years little has changed. I will argue
later that brain genes continue to evolve in response to changing social
conditions, which add in subtle ways to the repertoire of human behaviors.
For now, I merely claim that behaviors which are uniquely human, and which
are recently evolved, are most vulnerable to disruption by the appearance
of new selective forces.
If a new adaptation
has been selected for strongly, it might acquire robustness even in a relatively
short time. Human language, which may have appeared 200,000 years ago, is
a candidate example. Language played such a crucial role during its evolution
that the genes that code for it are probably already robust.
The capacities for
reading and writing have a briefer evolutionary history, and the genes that
code for these abilities are more vulnerable. Until recently, few people engaged
in reading and writing. These genes provided a niche to only a small fraction
of the population during the past 4000 years. It is therefore not surprising
that dyslexia affects several percent of the population, whereas verbal language
impairment is virtually unknown.
Unintended Deleterious
Effects
I suffer from occasional
20‑minute blind spells, called "scintillating scotoma." It is an impairment
produced by a gene that in women produce migraine headaches. As I type with
difficulty through a flashing zig‑zag blind‑spell pattern, it occurs to me
that I am paying a penalty for some genetic mutation that is doing good somewhere else. Every mutation
does many small bad things for every big good one, and the sum of bad ones
found in most people must be worth their penalty; otherwise the gene allele
would not have evolved.
In the case of my
blind‑spells a dilated blood vessel is putting pressure on a nerve fiber carrying
signals from my eyes to my brain's occipital lobe. What if the dilation occurred
elsewhere within my brain? I might not know that it was occurring since I
could not see it. But it might nevertheless have subtle effects upon mood
or thought. There must be people, probably many people, who do indeed experience
mild mental afflictions, lasting 20 minutes for example, which are counterparts
to my scintillating scotoma. We should be prepared, then, for the possibility
that a certain amount of irrational human behavior is caused by genes that
are conferring a greater adaptation benefit in some other behavioral realm,
with the unintended side effect that behavior is mildly irrational in a different
realm.
I frequently think
about the penalties that are paid when evolutionary pressures for one trait
rise above the others. Sure, you can quickly evolve skin color in response
to latitude migrations, but you'll pay with other unintended defects that
accumulate, until the new skin has been achieved and a better balance of
evolutionary forces has been established.
Only 12,000 years
ago, just after the climate warmed but before the glaciers had melted enough
to raise the world's sea level, people in Siberia migrated across Beringia
to the new world. As they moved south, generation after generation they would
have lost their need for light skin. Central American Indians are dark‑skinned,
and this must have been achieved in less than 10,000 years. But those who
continued their migration southward, past the equator, they would have needed
to re‑achieve light skin. Perhaps at each migration juncture those who were
best adapted to the latitude stayed behind and the others continued the
migration. This could have minimized the risks of unintended deleterious
mutations, but it is more likely that the southward migration was so hurried
that skin color played no role.
Such fast adaptations
must have produced defects in other aspects of the American Indian. Perhaps
they lost the ability to metabolize alcohol; we shall probably never know
what compromises the genes had to make to adapt quickly to the need for a
different skin color.
Cancer may afflict
humans more than most other species because we have recently undergone a
rapid evolution under strong selective forces that rewarded brain re‑wiring
(to accommodate behavioral adaptations) and immune system enhancements (to
fight pathogens seizing the opportunities offered them by the newly evolved
super‑tribe human lifestyle). To achieve these new traits, genes must have
been selected that would normally not be acceptable because of their unintended
deleterious effects, and a defense against cancer may have been one such compromised
ability.
The Dangers of Fast
Evolution
Species evolve at
different rates. Even a given species may remain genetically static many generations,
then respond to an abrupt change in climate by
evolving fast. Rates of change must vary by orders of magnitude, with long
eras of equilibrium punctuated by short periods of disruptive change. Mammals
lived throughout most of the dinosaur era, and flourished only after the
meteorite impact of 65 million years ago (which killed the dinosaurs because
of a brief, disruptive climate change lasting several years).
The equilibrium
periods are available for "clean up" of unintended deleterious effects created
during the fast evolving times.
The great diversity
of human anatomy, relative to other animals, testifies to the great potential
for fast human evolution. Strong selective forces must have superceded such
things as head shape, for example.
When a species is
suddenly subjected to a strong selective pressure, a few gene sites will suddenly
grow in importance. More than two alleles may exist at each "hot" site (if
only one allele exists, it won't be a site for selective pressure). Other
sites, being relegated to lesser importance, are likely to accumulate mildly
deleterious mutations with less consequence than before the fast evolution
(to use a metaphor, it's as if no one is "minding the store" when a new one
appears). Humans, who have been evolving fast for the past 7 million years
(since separating from the chimp lineage) must
have many multi‑allelic gene sites. The more alleles
that are in competition, the greater the fraction of maladaptive offspring.
Thus, the faster evolution occurs in response to some new selective pressure,
the greater the likelihood of a low offspring survival rate in order to prevent
a proliferation of the unfit.
Is it not ironic
that today, after coming out of a phase of extremely fast evolution in several
traits, humans have just achieved what must be the highest offspring survival
rate ever? Does this not mean that humans also must be exhibiting the greatest
rate of survival of maladaptive individuals? How long can this last? This
topic will be returned to in Chapters 6 and 8.
Lag and Regression
An abrupt environmental
change, such as those at the onset of an interglacial (occurring every 120,000
years, typically), must set evolution in motion in new directions. Until
a new "optimum" has evolved, producing stasis (and genetic consolidation),
there will be "lag." Some things are easier to evolve than others, and they
will lag less. Skin color may be one example.
Because adaptation
takes time, there could be a lag in many traits after an environmental change.
Present aspects of human nature should "make sense" only in the Pleistocene
context, not necessarily in that of the Holocene (the past 12,000 years).
For this purpose it has been useful to create the term "environment of evolutionary
adaptation" (EEA), also referred to as the Ancestral Environment (AE). Common behaviors that were adaptive 20,000 years ago need
not be "adaptive" today (Symons, 1979).
The Yanomamo Indians
of South America appear to be more "primitive" than their Asian stock who began migrating to the New World ~12,000 years ago.
How can this happen? Could the forces of evolution actually cause a population
to regress? Yes! And maybe this happened with the Yanomamo. Their regression
is only in relation to what was adaptive in their former setting. By definition,
they must be better adapted to the Venezuelan jungle than were the original
Siberian stock, or even the partially modified Central American Indian stock.
The longer the race,
the greater the disparity between the contestants – especially between winners
and losers. This is certainly
true for a foot race, but is it true in evolution? Consider that our ancestry
traces back to a chimpanzee‑like animal 7 million years ago, or 2 billion
years ago to a one‑celled life form, and 3 or
4 billion years ago to strands of DNA. Things like those early DNA strands
may exist today, as do many one‑celled life forms that may resemble those
in our ancestry. So "yes," the longer the race, the greater
the spread between the evolutionary contestants (note that all extant living
forms are "winners").
In human affairs
there is a discernible spreading in the quality of life of winners and losers.
The most prosperous people of today have a higher standard of living than
the most prosperous of yesterday, yet there are people living today who are
no better off than the worst off yesterday. Can there be stability in a
world where the rich get richer, and the poor stay poor? This is a topic
for Chapters 11, 14 and 16.
Evolutionary Reversal
Random mutations
rarely produce benefits to the individual organism (i.e.,
for the ability of the organism to stay alive, out‑compete its contemporaries
and to out-reproduce them). A mutation that alters a gene is likely to have
effects on many phenotypic traits (pleiotropy), and usually all or most traits
suffer from random mutations. For a mutation to succeed, it must confer
some advantage that outweighs damage done to many
other traits. "Forward" evolutionary change is thus difficult.
After a genetic
mutation spreads throughout a gene pool, it becomes part of a genetic setting
that new mutations must deal with. If a new mutation relies upon the presence
of the first one, and if this second mutation also spreads throughout the
gene pool, then the first gene has a more secure future. This occurs because
any challenge to the first gene must confer an advantage that outweighs the
contributions of two genes ‑ the first one and the other gene that relies
upon the first one for it's proper expression.
The longer a gene stays within a genome the greater is the chance that other
genes will become dependent on it and therefore provide it with additional
security. When this happens, the gene has become "entrenched."
Consider the situation
of environmental change that reverses itself at some later time. The first change may lead to the appearance and widespread
acceptance into the genome of a mutation. Let us assume that this new gene,
which has almost completely displaced an older one, confers an adaptive advantage
in the new climate. Suppose, now, that before this new allele has time to
become entrenched, the climate changes back to the original state. The few
individuals who carry copies of the original gene allele will become a source
for the quick re‑emergence of the original allele. Evolution can be said
to have "reversed" itself.
If the second climate
change occurred much later, however, this evolutionary reversal might not
be feasible. First, the original allele may have disappeared, and second,
other genes may have become dependent upon the presence of the new allele,
making it more difficult to dislodge from its entrenched location. In theory,
both difficulties for an evolutionary reversal can be overcome, but they
may constitute an insurmountable obstacle to the reversal.
Laboratory evidence
exists for "reverse evolution" (Teotonio and Rose, 2000). Fruit flies from
a standard stock were selected for various experiments over the course of
20 years (200 generations) and were subjected to new environments to produce
variant strains. When fruit flies from these new strains were subjected to
the original environment, in every case reverse evolution was observed. In
two cases, the reversal was almost complete after only 10 generations; others
required 50 generations. In some cases the amount of reverse evolution was
small.
At every instant
of a species evolutionary history, the most vulnerable genes are the most
recently‑acquired ones. This concept will be returned to in later chapters.
Culture can be thought
of as a collection of "culturgens" or "memes" ‑ similar to a genome being
comprised of a collection of genes (Lumsden and Wilson, 1981). Although some similarities exist between genetic and cultural
evolution, the differences are striking. This topic will also be dealt with
in a future chapter (Chapter 16), as a unifying theory for understanding
the rise and fall of civilizations.
Mutational Load
Although the idea
of "mutational load" was described by Kondrashov (1988), we owe H. G. Wells
it's first brief expression (ironically, in the
same journal, almost 100 years earlier). In 1895 Wells wrote: "Has anything
arisen to show ... that where the life and breeding of every individual of
a species is about equally secure, a degenerative
process must not inevitably supervene?" (Wells, 1895).
Primitive people
today produce about 7 offspring per woman. Allowing for slightly shorter
life spans in past times, about 6 offspring per woman was normal. On average,
only 2 survived to adulthood. Is it possible that some of the 4 who died were
genetically inferior? Yes, of course.
Approximately half
of all conceptions fail to produce a live birth. It is speculated that the
half that die are genetically defective due to some incompatibility between
the paternal and maternal alleles. It is a small step to suggest that there
will be a residue of live births that are also destined to fail to survive
childhood due to genetic defects. If this is true, then what would be the
genetic consequences of intervening medically to sustain all live births
through childhood and into adulthood?
If some of the 2/3
of live births that formerly died were due to genetic defects (a fraction
derived from the ratio of childhood mortality rates in primitive and modern
societies), and if all live births now live a full and reproductive life,
then surely the genetic defects which they carry will be contributed to the
gene pool in larger numbers than would have occurred in the ancestral environment
(AE). Our gene pool must inevitably accumulate these defective genes at
a higher rate than in the past. This phenomenon is
called "genetic load" (Kondrashov, 1988).
It may be impossible
for a species to average only one offspring per adult for a long time. With
no excess of births, the downward pulling force of “genetic load” would degrade
the gene pool of the species. Therefore, the survival ratio must be kept
well below one if humanity is to maintain a healthy genetic future! We who
survive without serious genetic defects should be grateful to those less
fortunate, whose deaths in the past made us possible.
I feel sorry for
the bent masses of future people, for they will suffer from cruel disabilities
that were traditionally weeded out by the neglect of less benign times in
the AE. Humanity reaps what it sows, and it is sowing the wrong genes ever
more often and preserving defective offspring with an excess of unthinking
compassion.
Compassion can be
a double‑edged sword. What seems laudable for one generation may in fact create
unlaudable consequences for many future generations. I shall return to this
moral dilemma in Chapter 11.
─────────────────────────────────
CHAPTER
5
─────────────────────────────────
GENETICS TUTORIAL
‑ PART III
Adapting to Novel Environments
I want to distinguish between
"outlaw genes" and those that are innocent by virtue of a changed environment.
The genes that reward eating sweets are usually mal‑adaptive in today's setting,
but in the ancestral environment (AE) they were adaptive. If we moderns
lived in the AE we would categorize the sweet‑tooth genes as helpful to
individual welfare as well as genetic welfare. This concept can be illustrated
using something that is well known in the remote sensing field called statistical
retrieval theory. This is described in Appendix C; a very brief description
is given here.
Figure 5.01 The filled square region
contains environments, physical and social, that have been encountered in
the past by our ancestors, and for which we are adapted.
The open square symbols represent modern world environments which humans
are experiencing for the first time, and for which we may not be pre‑adapted.
This is a mere 2‑D representation of a many‑dimensioned world environment.
The basic concept is that evolution
creates organisms that are relatively well-adapted to their AE, and when
the environment abruptly changes to something that the species has never
experienced the individuals are likely to be mal-adapted in ways that cannot
be easily imagined. In theory the individuals could be better off in the
new environment, but it is more likely that they will be worse off.
In the above figure, think
of the region in which the solid square symbols are found as corresponding
to one climate regime, such as a jungle environment, and think of the open
squares outside that region as representing the environmental conditions
for another climate regime, such as the glacier's edge. When our human ancestors
migrated northward from Africa into Europe they were moving from one
environment to another, and some of their genes had to “adapt” (be replaced).
One of the genes that adapted controlled skin pigmentation. Others were nose
length, eye color, hairiness, stature, etc. After the adaptations were essentially
accomplished, the new set of genes achieved a better performance in the
new reality space. It could be said that the new race of people were comfortably
within the environmental regime indicated by the dotted circle and solid
squares in the above figure.
Adapting to Changeable Environments
There is also the matter of
having to adapt to a climate that has greater annual variations. At mid‑
and high‑latitudes the seasons are more pronounced than in the tropics; a
tribe of people who must endure climate extremes during the course of a year
will have to adapt to a wider range of conditions than a tribe that lives
in the tropics. Cro-Magnon man, who evolved adaptations for the mid‑latitude
climate extremes in Europe, must have made the compromises
that rendered him less than perfectly adapted to hot summers and cold winters,
yet able to survive in both. A region that is subject to periodic climate
changes, which occur faster than the time needed for the gene pool to evolve
a new adaptation, is in effect a region with a bigger reality space to which
the genes must adapt.
For another example, El Nino
weather patterns repeat every 4 to 7 years, creating at some mid‑latitude
regions shifting amounts of rain, temperature and other seasonal properties.
It is unclear how long El Nino/La Nina cycles have been occurring, but this
is a convenient example for illustrating how our ancestors who had left the
jungle may have had to deal with wide ranges of reality space. When the
genes adapt to climates that shift back and forth on timescales that are
shorter than evolution can track, the adaptation will have to be for the
entire range of climates and fauna. However, if the range of settings is
large, penalties will grow for life within each setting.
Tolerating Diversity as a Solution
One way the genes may have
solved this problem is to "tolerate diversity." In any diverse population
some individuals are likely to be pre‑adapted to never‑before encountered
environments. This is a "group selection" argument. Populations that are
relatively isolated compete without coming in contact by merely surviving
or perishing when environmental conditions change wildly. We can speculate
that those that survived will be the ones that tolerated diversity, given
that some of their members were pre‑adapted for future conditions. Such populations
would be especially pre‑adapted for climate changes that had never occurred
in the past. The drastic climate fluctuations that occurred during the transition
from Pleistocene to Holocene (18,000 to 10,000 years ago) would have been
relatively unprecedented (a similar period of climate change occurred at
about 120,000 years ago). Thus, the introduction to the Holocene may have
favored those tribes that were inherently more tolerant of diversity.
Evolution of
Behavior Repertoires
Most environmental changes
are repetitive, such as the El Nino/La Nina cycles. Environments
that occur at intervals of less than 10,000 years, for example, are candidates
for another genetic solution, described next.
When an environment changes
wildly a person may take a reading of present conditions, which could be
climate, population density, food scarcity, or social setting, and then change
behavior in response to the perceived change of setting. Humans have a larger
repertoire of behavioral responses to situations than any other animal! Humans
whose ancestors have encountered a variety of distinctly different environments
may have unknowingly prepared their descendants for a faster “within a lifetime”
adaptation to any of these environments compared with humans who have never
encountered the same range of environments. This is asking a lot from natural
selection, for we are assuming the creation of individuals who are pre‑adapted
in a very sophisticated way to environmental change. These people are capable
of instinctively responding to a specific environmental change by changing
their behaviors in a specific manner that is adaptive. Is it asking too much
to invoke the evolution of this capability?
In essence, we're asking if
natural selection can evolve a human brain that has circuits that do the
following: "IF (this setting) THEN (employ that behavior
or lifestyle)." These circuits are analogous to the human immune system's
large repertoire for doing "IF (this pathogen) THEN (employ that immune response),"
as pointed out by Gazzaniga (1997). We know that the human immune system is
immense, so the evolution of the capability is apparently possible. Its evolution
may have been forced by the coming together of tribes to form large settlements,
and eventually urban centers. Some diseases flourish when population density
is high, or when the population size is large. These new diseases would reward
people with more capable immune systems.
I am suggesting that humans
today are prepared to read their setting and shift their behaviors, and even
their group's lifestyle, in a way that is adaptive. An extreme example would
be a tribe that is sedentary when the environment produces abundant supplies
of food, but switches to a hunter/gatherer mode when the environment is
less bountiful. When the switch occurs, requiring a new lifestyle, many things
related to behavior might have to change ‑ such as marriage customs, property
ownership, status hierarchies, etc. The genes would simply code for a switch‑over
in many behaviors in response to a new perceived setting.
Our ancestors probably encountered
many environmental changes, especially during the Holocene, presenting many
opportunities for the genes to develop a reliance on requiring lifestyle
mode changes based on a perception of "conditions." The adaptation to variable
environments would simultaneously have rendered us physically adapted to
no one environment in particular, giving us the appearance of inferiority
to other animals, which are well adapted to narrower range of environments. The human brain, on the other hand, has become
capable of switching between a large repertoire
of behaviors, and when a mode switch is made correctly, the new lifestyle
can be well adapted to the new environment. These factors are ideal for the
creation of "culture" ‑ which allows for quick behavioral "adaptations"
to environmental changes.
How lucky for humans if a fluctuating
environment produced mental abilities for adjusting behavior that were made
available to the challenges of non-environmental changes. It must be common
for a mental “tool” to be created in response to one challenge and only later
become useful for other tasks.
Risks of Behavior
Repertoires
How unlucky for humans if this
same capability for achieving adaptive changes in lifestyle by “taking readings”
of one’s setting could render civilizations vulnerable to “opportunist” individuals.
This speculation is dealt with in Chapter 11, 14 and 16.
The mismatch between the modern
brain, evolved for an ancestral environment, and the modern world, recently
shaped by Man himself, is treated (but not from a rigorous sociobiological
perspective) in the book New World, New Mind: Moving
Toward Conscious Evolution, by Ornstein and Ehrlich (1989).
Later chapters will come back
to this point, so for now just remember that the modern world is a man‑made
environment with very little of the ancestral environment to provide assurance
that our living in it will appear to be adaptive, or even stable.
─────────────────────────────────
CHAPTER
6
─────────────────────────────────
EVOLUTION CONCEPTS
AND HUMANS
"In a very real
sense human beings are machines constructed by the nucleic acids to arrange
for the efficient replication of more nucleic acids. ... We are, in a way,
temporary ambulatory repositories for our nucleic acids." Carl Sagan, The Cosmic Connection, Garden City, NY: Anchor Press,
1973.
Having described
some basic tenets of genetics in the previous two chapters, we are now ready
to undertake theoretical understandings of human behavior. It is important
to keep in mind throughout the rest of this book that all behavior is the
outcome of a competition among gene alleles for representation at specific
locations on chromosomes. Thus, the "macro‑behaviors"
at the individual level, which will be the subject of the rest of this
book, are the result of "micro‑motives" at the gene level.
GEP
An individual's
"phenotype" is "the way it is" ‑ its anatomy, physiology and behavior. An
individual's "genotype" (inherited genes) interacts with environment to
produce the "phenotype." Thus, a person's phenotype is who the person has
become, as opposed to who they might have become had their environment been
different. This powerful concept (Symons, 1979) can be referred to by the
equation: G + E = P, or GEP.
Anatomy includes,
for example, stature, the height a person achieves in adulthood. In a society
where food is plentiful, as for most people in the United States, stature is determined
almost entirely by the genes. But in a society where food is scarce for
some people and plentiful for others, stature is determined most strongly
by food availability (the environment, unless the genes determine access
to food).
Consider the case
of Japanese stature before and after World War II. Children after the war
grew taller than their parents. Between the generations the disparity in
the availability of food was large, and this diet difference was large "in
relation to" the genetic variation between individuals. Stature differences
were determined almost entirely by environment. This example illustrates that
if the variation of environment is large, environment can be the dominate
cause of phenotype variation, whereas if the variation of environment is small,
making the variation of genotype more important, genotype can be the dominate
cause of phenotype variation.
Physiology includes,
for example, immune response. The genes create an immune system that includes
a repertoire of responses to specific pathogen stimulations. A virus will
elicit an immune response that is appropriate if our ancestors were survivors
of the same (or similarly‑shaped) virus. The repertoire is limited by ancestor
experience, so an individual is likely to be vulnerable to new viruses
with novel shapes.
If the environment
harbors the same viruses and bacteria that our ancestors survived, and if
we are considering a stable population with no immigrants from other regions,
then essentially all people who get sick will recover, and it will not be
apparent that genotype is affecting phenotype (vulnerability to disease).
But if the population includes immigrants from distant places, where there
has been a different virus exposure history, there may be dramatic differences
in who recovers and who dies from local sicknesses. The immigrants will be
at a disadvantage when infected by local viruses and the native population
will be vulnerable to any viruses brought by the immigrants. For dramatic
illustration, old world explorers came to the new world and brought diseases
that killed most people in the new world (Diamond, 1996).
The "behavior" component
of phenotype is the most interesting, and the most challenging to understand.
The "immune system's response repertoire" is a useful starting analogy for
understanding behavior (Gazzaniga, 1992; Gazzaniga, 1997; Jerne, 1967). A
"stimulus" in the environment can produce a "behavioral response," as when
an abrupt approach of something to the face produces an eye blink. We blink
our eye because we have ancestors who survived more successfully than those
who didn't have the eye blink response.
A reductionist will
want to employ the "stimulus/response" (S/R) explanation for behavior as
much as possible. With effort, this approach at understanding behavior is
broadly successful more often than is conventionally acknowledged. Since it
is the simplest possible explanation type, it should be invoked as a first
hypothesis.
S/R fails to account
for behaviors that are self‑initiated, i.e., motivated behaviors.
For example, this morning I decided to hike in the mountains in the afternoon.
Planning a day's activities, as with life goals, requires something that
in humans is identified as "prefrontal" cerebral activity, and a general sub‑cortical
"drive" mediated by a "reticular activating system" (as described in Chapters
7 and 8). The prefrontal cortex initiates broad goals, such as a career path,
it initiates behavioral programs, such as preparing a speech, and it also
initiates specific behaviors, such as talking.
I will argue that
the "shape" of behavioral programs, and the "shape" of life paths, are initiated by brain circuits that we inherit. The
specific behavioral programs, and specific life paths, are the product
of an interaction between inherited brain circuits and the environment.
Not only is an individual's
environment a changing thing (career opportunities, available books, current
beliefs, etc), but the human environment can change dramatically from one
generation to the next, and especially from one millennium to another. It
is very likely that everyone was capable at birth of adapting to a hunter‑gatherer
lifestyle, as was common in the AE (ancestral environment).
A human trait that
seems to be common, such as "greed," may be expressed in only specific environments.
A person born into a tribal hunter‑gatherer setting, where there are few
possessions (because it's hard to carry things from camp to camp) may grow
up without expressing greed. The same person growing up in an agrarian society
might be greedy. In other words, the person's "genotype" will interact with
"environment" to produce a lesser or greater amount of "greed" (phenotype),
which illustrates G + E = P.
There are limits
to an environment's influence. This is easiest to illustrate using dramatically
different genotypes, such as individuals of different species. We cannot
make a snake behave like a cat just by cuddling it while it grows up. The
snake is limited in what it can become. No amount of environmental adjustment
will ever make a snake cat-like, because snake genes do not have cat behavior
in their repertoire. Instead of Gsnake + Ecat = Pcat we are limited to Gsnake
+ Ecat = Psnake. Although snakes and cats are different species, the GEP
equation still holds. Illustrating this concept using such different critters
the point is easier to understand.
In any population
there is a variation of genetic predispositions, or genotype. Thus, given
a fixed environment, there will still be a variation in phenotypes, and in
this case it will be due entirely to the variation of genotypes. Where there
is wide variation in the environment, even a uniform genotype will produce
a variation of phenotype, and such variation will be due entirely to environment.
The normal situation, of course, is for variations in both genotype and
environment, which obscures the sources of observed phenotype variation.
Every population
must have unfortunate cases of bad genotype coupled with bad environment.
Whereas either one might produce a bad adult, together
they could produce a really bad adult. (Do you think Attilla the Hun
may have been the product of bad genes and a bad environment? Was he maladapted
from the perspective of his genes?)
IF/THEN Brain Circuits
It is reasonable
to assume that each person inherits genes that pre‑wire their brain to recognize
situations that elicit appropriate behaviors (S/R) for situations that our
ancestors repeatedly encountered and survived. Like any computer program
with many IF/THEN sections of code, some of the IF/THEN code will not be used
during normal experience. Indeed, most IF/THEN code that exists may only
be used in response to rare experiences, especially so for humans, who have
an unusually large repertoire of conditional behaviors and personality development
paths.
Once a specific
piece of IF/THEN code comes into existence, in response to a sustained period
of selective pressure in which a recognizable situation occurs and responses
have reliable benefits or costs, this piece of code can remain in the genome
almost forever. If it later comes into conflict with a similar situation
requiring different responses, then this old code will be modified or may
disappear. Since the code that elicits a behavior is almost always produced
by a combination of genes, if any of these genes are modified in response
to other adaptive pressures the original code could inadvertently be modified.
If this occurs, the gene pool would have to be exposed to the original adaptive
pressures again to restore the original, or equivalent, IF/THEN code.
If it someday becomes
possible to list the IF/THEN circuits in a typical human brain, we may wonder
when and where each evolved. If this were possible it would probably turn
out that most code sections were created during specific eras in our ancestry,
with few (or no) recurrences. Each distaste may
thus owe its existence to a time when we lived among a specific inedible
plant. The plant might have existed for only a few centuries, during the past
2.5 million years of the human past, yet its IF/THEN legacy stays with us.
Many of our ancestors
were nomads, living in wandering tribes. Behaviors required by nomadic
tribal life are part of our repertoire, and if a "modern" non‑nomad were
raised in nomadic setting he might develop with the same nomadic functionality
as present‑day nomads. Settled farmers, living as single family units, are
also part of our ancestry, probably confined to parts of the past 10,000
years. Each of us probably has the code necessary to grow up into fully functional,
single‑family farmer. Large settlement living must have been a part of some
other of periods of our ancestry, confined perhaps to the past 5000 years.
Each of us is presumably capable of becoming functional urban dwellers. Our
large brains are "ready" for many lives that cannot all be lived!
Men Bear More of
Evolution's Burden
Paternity success,
as measured by offspring per male, exhibits a wider range than maternity
success (offspring per female). Every tribe will have some males who don't
reproduce, whereas it is rare to find women who are childless. For sexually
mature women, after weaning an offspring she is likely to become pregnant
soon after menstrual cycles return.
As with any species,
whenever a dominant male controls access of other males to females, there
will be a large disparity in breeding success among males. Harems were common
in human history, and presumably pre‑history as well. Even when males do
not dominate other males, females are prone to prefer to breed with specific
males.
Whereas women typically
give birth to about 6 or 7 babies during their lifetime, with little difference
between women, men may sire from zero to hundreds of babies! Why is there
such a disparity?
The human ancestral
environment is presumed to have been exclusively tribal. The men of most
tribes, it is thought, engaged in hunting expeditions. It was also probable
that they engaged in brief raids of neighboring tribes, as well as more dangerous
inter‑tribal warfare. Such male activities entail an extra burden of mortality.
A man could die not only from combat, but from a mismatch of anatomy or physiology
to climate. A man could also die by formulating less successful strategies
in warfare, or by not adhering to a planned strategy requiring careful social
coordination.
An extreme view
of this situation is to state that the purpose for men is to go out and be
measured. Those who come back, and especially those who come back as heroes,
will have survived the measurement test, and the women shall deem them more
valuable as potential fathers for their children. All women will prefer
to mate with heroes than with the others. (It goes without saying that they
won't mate with those who died in the process of being measured.)
Any man who refused
to "play this game," to go out and be measured, could expect to be shunned
by women ‑ as well as by men. Such a man may even have been banished from
the tribe (leading eventually to death). It has only been during the present
interglacial (during the past 12,000 years) that alternative niches proliferated
for the less adventurous man. Although there might have been a small, exempt
class of weapon makers, most men could not have escaped the high‑mortality
life style.
Consider the first
people who migrated from Africa to the mid‑latitudes.
Dark‑skinned men would have fared less well, all other things being equal,
than slightly lighter‑skinned men, as the dark‑skinned men would not synthesize
as much Vitamin D. Vitamin D deficient men would be at higher risk of succumbing
to the physical demands of traveling, hunting and warfare. On average, the
lighter‑skinned men would be more likely to return from exploits. The women,
who stayed home, would be less affected by lowered Vitamin D, so their mortality
would have been less affected than their male counterparts. After no more than 200 to 400 generations (based on the New World immigration experience),
the entire group's skin would have evolved to a new, more adaptive color. This process would
have been achieved by a differential survival of men, combined with a differential
breeding success of men. Thus, the burden of adapting would have been borne
more heavily by men.
Takeover Infanticidal
Males
Male lions kill
a female's young lion cubs after they overpower the male lions in a pride.
Not only does this remove lion competitors for the male's offspring, but
the female soon stops nursing, becomes fertile, and is available for mating
with the killer male. We humans might think that a female lion would be upset
to see her cubs killed by the new males, but amazingly, the female quickly
makes the best of a bad situation by becoming coquettish with the killers.
By these actions, the female increases the prevalence of the very genes that
thwarted her initial reproductive investment; by favoring this behavior, which
humans find so repugnant, the lioness helps shape the male genotype.
Infanticide by males
has been documented for species of birds, fish, insects and mammals ‑ such
as rodents, carnivores and primates (Wrangham and Peterson, 1996). The following
description is for primates.
"Hrdy noticed an
invading male charge after a mother, attempting to snatch away her baby.
For several days, the other females in the group tried to defend the mother
and her baby. But the male persevered, and finally managed to deliver a slash
to the infant's stomach that left the intestines exposed. Taking the wounded
infant to her breast, the mother looked up at the sky, as though in despair.
'It was the only time in my professional career that I wept.' // Because females are usually
outmatched in the physical war between the sexes, they are helpless to protect
their offspring against an infanticidal male. // Female gorillas respond to infanticide ... they
leave the father who allowed their baby to be killed and run off with the
murderous male. Infanticide, along with various female defenses, has been
seen in 13 primate species. (Angier, 1983.)
We humans find such
behaviors repugnant, so surely men do not act this way. Alas, they do! Studies of infanticide in Canada (Daly and Wilson,
1988a) and the United States (Daly and Wilson,
1988b) reveal that step‑fathers are 75 and 100 times more likely to commit
infanticide than are biological fathers.
In any species where
males "take over" a female by overpowering the resident male and kill the
offspring, females should evolve an aversion to males who cannot protect her
and her offspring from takeover males. Thus, women should find weak and low‑status
males unattractive, in relation to strong and high‑status males. This should
be especially true for attractive women, who are more likely targets for
takeover males. These predictions are borne out by "common knowledge."
Monogamy and Cuckolding
Monogamy, and the
associated female faithfulness which monogamous husbands require, give every
man a more or less equal influence on the next generation's genetic pool.
This must retard the potential speed of evolutionary adaptation of that gene
pool. Thus, the stronger the forces of evolution, the
greater the reward for polygamy.
It would be surprising
if the genes have remained blind to this. Women, for example, should sense
when evolutionary forces are strong, and in response, they should seek consort
with men who are "successful." If monogamy were the norm
(which would have been more likely only during the past 12,000 years), then
women should be expected to try to cuckold their husband (secretly mate
with a man who is not the husband) in order to bear children carrying the
more "adapted" man's genes. Since monogamy was probably rare before
12,000 years ago, in the human ancestral environment the need to cuckold
was also probably unimportant before that time. Cuckolding, I suggest, is
therefore a "recent" tool in women's behavioral repertoire.
Blood tests of Canadian
and American families reveal a cuckoldry rate that ranges from 15% to 25%
(see Christenfeld and Hill, 1995 for additional material). Presumably, cuckoldry
rate varies with time and conditions in accordance with some optimizing algorithm
created during the AE.
Knowing the optimal
time for a wife to cuckold her husband would have evolved during the time
that societies became monogamous. Refinements in a woman's cuckolding wisdom
would have improved the most when evolutionary forces were greatest. If a
gene pool underwent a period of polygamous evolution, the previously gained
cuckolding wisdom would have remained "ready" but not expressed until monogamy
was restored. This is similar to the way an immune system accumulates a repertoire
of immune responses, each specific one of which remains "ready" for expression
when exposed to a pathogen that is "recognized."
Recent studies (Hazelton,
2006) show that women have a heightened interest in cuckolding their husbands
when they are most fertile. This logical female strategy is matched by an
equally logical male strategy of exhibiting a higher level of mate-guarding
at the same time. The real challenge for women is to recognize when it is
appropriate to cuckold, and with whom. As for “when,”
being in a monogamous relation is one precondition.
Sensing that "times are tough" would be another (i.e.,
evolutionary forces are strong).
As for “with whom”
a woman must be capable of measuring her husband against other men. One measure
of the successful man is "fashion"; the man who is sought by other women
is likely to produce boys who will grow up to also be sought by the next
generation of women ‑ regardless of the intrinsic worth of the type. Another
way to identify a good candidate is to determine who is dominant over whom.
Men live much of their life within a male society, and the men who are most
successful in male activities, such as the cooperative hunt, will be accorded
privileged positions by other men. Women are sure to notice how men sort
themselves while establishing the male hierarchy,
and those who are esteemed by other men are good candidates for a cuckolding
episode. Women who are fascinated by men’s sporting events might be “doing
their homework” for optimizing their future cuckolding.
The main effect
of this increased attention by women to male worth is to increase the imbalance
of reproductive activity among men; fewer men will account for a greater
fraction of a generation's paternity. A secondary effect of this enhanced
reward for whatever the forces of evolution deem important is to reduce genetic
diversity. In a one harem society all offspring will resemble the harem master.
If he is vulnerable to a specific disease, most children of the next generation
will be similarly vulnerable.
Thus, there are
risks to tribal organizations that give excessive reproductive rewards to
small numbers of men. It is in a woman's genetic interest to not succumb
totally to fashion; but it is always in a man's interests to be the most successful
man and to dominate male reproductive activity.
Women understand
that husbands should be loyal "producers" even though they should favor other men for cuckolding. There are "husband material" men,
and then there are "exciting affair" men. Women are attracted to both types,
but in different ways. A faithful husband type is attractive at the time
a commitment is to be made, and the exciting affair type is less attractive
at this time. Some time after marriage, however, women's interest in "affair
men" should increase. Thus, women regard some men as good on long timescales,
serving as loyal husbands and fathers, whereas some men are good for short
timescales, serving as cuckold consorts to provide offspring with genes specialized
in victimizing the next generation.
Men likewise automatically
categorize women as good for the long term, serving as loyal wives and mothers,
while other women are good for short‑term consort. Men and women must automatically
categorize each other as belonging to one or the other category.
Men and Women Shape
Each Other
Men and women have
made each other what they are!
Men have a greater
variance in IQ, and we men also exhibit higher incidences of genetic deficiencies.
For example, dyslexia (reading and writing problems) is most common among
boys. This may merely be an effect of males exhibiting a greater genetic
variance of recently‑evolved traits (i.e., men dominate both
ends of the spectrum of most measures). Thus, there are more men geniuses,
as well as more men among the learning disabled. Men are burdened by "high
risk" mutation experiments that eventually benefit the larger population.
Men appear to be more "expendable" than women.
To what extent are
women responsible for making men genetically "fragile"? Women prefer men
who "go out to be measured," and who come back with good measures. When women
cuckold their husbands, they assure their male offspring a greater likelihood
of being a cuckolding partner in the next generation. These women also assure
that they will produce daughters who are prone to cuckolding their husbands.
This occurs because the cuckolding males are likely to carry genes which
predispose their girl offspring to cuckold when they are women, since they
are likely to have been the result of women who cuckolded (this is a subtle
argument).
Women shape men
with every preference they express. If women favor men who are "travelers"
(i.e., vagabonds, minstrels, pirates), then each succeeding generation of
men will tend to resemble travelers.
Why would women
be attracted to travelers? When diseases are a principal
cause of mortality, traveling men that women encounter are the ones who have
immune systems with the best immunity to diseases beyond the village. This
may account for girls going crazy for pirates, traveling musicians, and
other itinerate roustabouts who have no long‑term parenting value.
Birth Order
Frank Sulloway (1996)
has presented an immensely well documented case for the influence of family
birth order on specific personality traits. The theoretical argument for
such an influence begins with the fact that in the ancestral environment children
often died before reaching adulthood (approximately 2/3 of children perished).
Surviving childhood requires that the child adopt strategies for maximizing
parental investment. By this logic, firstborns should ingratiate themselves
to their parents, and gain their favor by appearing to be good prospects
for their investment. Firstborns should be obedient, conscientious, hard‑working,
and they should internalize the values of their parents.
Laterborns, noticing
that there already exist firstborns who have acquired parental confidence
by becoming what the parents want, must create for themselves a different
identity. They must distinguish themselves from their older sibling by excelling
in another endeavor, for if they tried to compete in the same arena, and
became equally successful at comparable age, they would be destined to always
be a worse investment prospect due to their age disadvantage. All other things
being equal, older children are a better investment option because they've
already survived more of the childhood risks, and they are closer to childbearing
age. Thus, laterborns try to excel in things untried by the firstborn, and
perhaps unfamiliar to the parents. Laterborns are more open to new experiences,
and are more adventurous. As stated by Sulloway (pg 98), "...the addition
that each child makes to the parents' inclusive fitness will tend to be proportional
to the development of skills not already represented among other family
members."
Firstborn boys followed
by a laterborn girl are a congenial combination, since the girl is naturally
inclined to have different interests than the boy. From the parents’ perspective,
both children are like first-borns and represent good investments.
Firstborn boys feel
threatened by a laterborn boy. They are likely to fight, and the younger brother
must become proficient with wit, words or some other clever strategy to compensate
for his smaller body. The older boy will become accustomed to dominating
his younger brother, whereas the younger brother will become adept in the
use of social skills for minimizing the disadvantages of being dominated
by the older brother. These effects appear to be maximum when the age difference
is about 3 years (close enough in age to be competing for similar age‑related
niches, yet different enough that the younger is weaker and can be successfully
dominated). Sulloway writes (pg 79) "Like the alpha males of primate societies,
firstborns covet status and power. They specialize in strategies designed
to subordinate rivals."
To the extent that
Sulloway's birth order correlations are correct, women should prefer men
who happened to be firstborns with younger brothers. They are more likely
to be "masculine" and capable of protecting their wives from "takeover" males.
On the other hand, these men are less likely to tolerate cuckolding wives,
rendering cuckoldry a more dangerous option for the wife of a firstborn husband.
How confusing (at
a subconscious level) this birth order "environmental monkey wrench" must
be to women! Men who appear to be strong and domineering may be so merely
because they had a brother 3 years younger. They can probably be counted
on to protect them and their children from takeover males, but they will not
necessarily provide "domineering" and "high status" genes to her offspring.
Do women have methods for identifying "genotype‑produced" versus "birth order‑produced"
dominant men? We await further studies in this young field.
Sexually Specific
Morality
Women complain that
men can philander with less consequence than women who cuckold. They attribute
this disparity to the fact that men can get away with dominating women due
to their greater physical strength.
The real reason
for this duality of morality has to do with the difference in natural consequences
for out‑of‑marriage mating. A philandering husband does not necessarily diminish
his "paternal investment" value for his wife's children, whereas a cuckolding
wife who produces illegitimate offspring necessarily does cause her husband's
"paternal investment" to be squandered.
If it Feels Good, Beware!
What is the purpose
of emotions? They are meant to influence behavior!
In the particular
case of humans, emotions are meant to influence behavior in situations where
rational thought is also likely to subvert the genetic agenda. Some behaviors
are too important to be meddled with by rationality. The Australian redback
spider, in which the male is prone to allow itself to be eaten by the female
during copulation, is incapable of rational thought. A simple, automatic
instinct suffices to assure that his gene‑serving deed be done. But what about humans?
Humans think, and
are theoretically subject to influence by rational thought. The genes, in
their infinite wisdom, have created emotions to safeguard behaviors that
serve their interests. Emotions are employed to protect behaviors that are
threatened by rational considerations of individual welfare!
Emotions symbolize
the conflict between "outlaw genes" and a thinking, rational individual.
Therefore, if something has a feel-good emotional payoff, beware!
This theory implies
that only intelligent creatures have emotions. It suggests that emotions
were "invented" by the genes as a quick solution to a fast evolving human
intellect. The genes could not know what rational threats lay ahead, in untried
environments, or even newly endowed rational brains, yet they "learned" from
past experience that certain actions were at risk of being overturned by individuals
who cared more for individual welfare than being an obedient tool of the
genes (of course the genes didn't "know" anything; it merely happened that
those that safeguarded important behaviors from the influence of other gene
mutations prospered.)
We will return to
this subject in a later chapter.
Consciousness
It is difficult,
at this point in my argument, to avoid the problem of "consciousness." It is tempting to speculate that C, as consciousness gurus
refer to it, was invented by the genes to mediate conflicts between an old
instinctual brain, and a new rational one. To the extent that C grew in power,
emotions must also grow in strength.
A common sense theory
for C is that it “exists” whenever a novel situation demands that brain modules
compete for control of understanding and behaving. C almost certainly is
generated by the prefrontal cortex, possibly on the left side. Measurements
of brain activity show that this area is active when a novel task is being
confronted, whereas tasks that have been mastered during previous encounters
are not associated with the same level of activation. These cortical activity
measurements may have been detecting something produced by C.
Are humans more
conscious than chimpanzees? There is growing evidence that chimpanzees "think"
‑ in the way that people commonly think of thinking (Goodall, 1986, and Wrangham
and Peterson, 1996). Chimpanzees appear to have something called “theory
of mind,” or knowing what other chimpanzees are likely to know, and this
also would imply that they have C, at some level.
It is important
that thinkers with a sociobiological approach address the consciousness problem,
as most of the C literature is devoid of an appreciation that 1) genes construct
brains, and 2) genes exist because they're good at surviving. Anyone else
who tries to investigate C is handicapped at the outset.
Books that treat
consciousness with an adequate respect for the reductionist paradigm include
Consciousness Explained (Dennett, 1991), The
Illusion of Consciousness (Wegner, 2002) and The Quest for
Consciousness (Koch, 2004).
In the next chapter
we will return to this issue, which deals with neuropsychology and evolution
─────────────────────────────────
CHAPTER
7
─────────────────────────────────
BRAIN ANATOMY AND
FUNCTION
“…intellect
arose merely to serve the will [genes]. Most men … are incapable of any other
employment of their intellect, because with them it is merely a tool in
service of their will and is entirely consumed by this service…” Schopenhauer, Aphorisms (1851).
"Men
think themselves free because they are conscious
of their volitions and desires, but are ignorant of the causes by which they
are led to wish and desire." Spinoza, Ethics (1677).
The brain is an
organ meant to help genes survive, and in this respect it is no different
from the heart, liver, and reproductive organs. A thinking brain may not
like this assessment, and it may prefer to view the body and its organs, as
well as the genes, as existing to serve the brain. But modern science, spearheaded
by sociobiological insights, is once again forcing Mankind to move further
down from his pedestal by discrediting another cherished belief. This chapter
will describe brain anatomy and function. The next chapter will address their
evolution.
Part of my intent
for this chapter is to remove some of the "mystery" from how the brain works.
I want to convey a sense that the brain functions like a "machine," and that
living things are automatons, consistent with this book's reductionist approach.
Brain Anatomy: Vertical,
Horizontal and Front/Back Layout
The human brain
consists of a primitive hindbrain, a small mid‑brain section, and a large
and complicated forebrain.
The hindbrain, which
began its evolutionary existence ½ billion years ago, resembles the entirety
of a reptile brain, and has been referred to as our "reptilian brain." The
hindbrain's "stem" connects to the body; it receives information from sense
receptors and issues commands to muscles and body glands via the spinal cord.
The hindbrain's cerebellum stores motor commands and produces smooth movements.
The mid‑brain has
a minuscule function, and won’t be described here.
The forebrain, on
the other hand, is where uniquely human attributes are generated. It includes
a limbic system, thalamus, basal ganglia, and two large cerebral hemispheres.
The limbic system has many components; it maintains homeostasis (body temperature,
heart rate, blood sugar, etc), and controls emotional state (things like
hunger, anger, fear and sexual arousal). The limbic system's pea‑sized hypothalamus
performs many of these functions using electrical commands, some of which
activate hormone producing glands in the brain. The thalamus and basal ganglia
control conscious state and initiate movement, respectively.
The cerebral cortex,
comprising 70% of human brain volume, consists of a left and right cerebral
hemisphere, with an interconnecting corpus callosum. Although the cerebral
cortex is only 1/8‑inch thick, its surface area is about 1 ½ square feet,
and it has evolved a folded configuration to allow the surface to fit within
the human skull. The inside surface of the cortex (gray matter) has an immense
number of nerve fibers (white matter) providing connections to other parts
of the cortex, the limbic system and other brain components.
The cortex is the
most recently evolved part of the brain, and fortunately it is also the most
accessible to study. The left cortex and the right cortex each consist of
4 lobes: occipital, parietal, temporal and frontal. The occipital "sees,"
the parietal "feels," the temporal "hears," and the "frontal" thinks and
commands! The "see/hear/feel" lobes are referred
to as "posterior lobes" (since they comprise the rear half). They can be
thought of as "receptive lobes" since they receive input from the body and
environment. The "see/hear" lobes receive "remote sensing"
information (visual and auditory input), while the "feel" lobe receives
in situ information (touch, temperature, pain and body part
position). The frontal lobes, the front half
of the brain, receives input from the posterior lobes, and they “think”
about the situation, formulate action plans and issue commands to muscles.
Figure 7.01. Brain lobes:
Frontal, Parietal, Temporal, Occipital. View is
of the left side, front is toward the left.
The corpus callosum
(not shown) connects all four lobes of one side to the corresponding lobes
on the other side. This nerve bundle is located underneath the frontal and
parietal lobes, at about the same level as the temporal lobe.
Primary, Secondary
and Tertiary Cortical Areas
Each of the 4 lobes,
the frontal, parietal, temporal and occipital, consists of 3 cortical areas:
primary, secondary and tertiary. These are shown in Fig. 7.02 using the numbers
1, 2 and 3.
The part of the
parietal lobe bordering the frontal lobe, area P1 in Fig. 7.02, is the "sensory
cortex" (or "somatic cortex"). This strip of cortex is where in situ sensory signals from the body arrive. Next to the somatic
cortex P1 is an area, F1, located in the frontal lobe and called the "motor
cortex" or "motor strip." The motor cortex issues commands for movement
(requests, actually, since sub‑cortical regions may "veto" the requests).
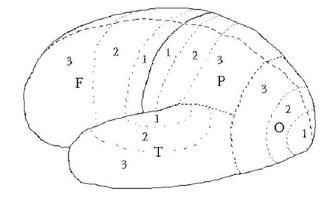
Figure 7.02. Approximate boundaries for cortical primary (1), secondary
(2) and tertiary (3) areas in each lobe.
There's a one‑to‑one
mapping of body location to position along the sensory cortex strip, P1,
and motor cortex strip, F1. Starting from the part of the strips closest to
the center‑line (top of brain) and going outward, body positions are allocated
in the following sequence: leg, neck, head, arm, elbow,
etc, to face, lips, teeth and tongue.
For the posterior
lobes raw sensory information arrives at the primary cortical areas, which
deliver processed versions to the secondary areas, which in turn deliver
even further processed versions to the tertiary areas. The 3 posterior lobe
tertiary areas border each other, and this is where the most "conceptualized"
versions of perceptions are inter-compared and elaborated.
Figure 7.03. Flow of nerve activity when something is "felt."
For example,
when something is "felt" the flow of nerve activity "flows" according to
the depiction of the figure above. When something is "heard"
the flow of nerve activity "flows" according to the depiction of the following
figure.
Figure 7.04. Flow of nerve activity when something is "heard."
When something is
"seen" the flow of nerve activity "flows" according to the depiction of the
following figure.
Figure 7.05.
Flow of nerve activity when something is "seen."
For each posterior
lobe the pattern of nerve activity is the same: primary activity leads to
secondary activity, which then leads to tertiary activity. The next step
is for tertiary activity in adjoining areas to “compare notes,” or interact
with each other.
Tertiary Cortex
Convergences
When a familiar
object is recognized a small set of tiny nerve circuits are set into "resonance."
For example, when a coffee cup is seen, there's a flow of activity in the
occipital lobe from primary to secondary to tertiary. When it reaches secondary
cortex, i.e., O2, there will be sub‑features such as handle,
rim, steam, etc "active" at their respective locations in O2 (created from
interaction with the environment in childhood). These interact in O3 (occipital
tertiary), setting into resonance a tiny circuit corresponding to "coffee
cup seen."
Figure 7.06. Nerve activity when a "coffee cup" is seen.
The same coffee
cup can be felt. In this case the nerve activity will be as shown in the
next figure.
Figure 7.07. Nerve activity when a "coffee cup" is felt.
The coffee cup may
be heard, as it is set down on a table. In this case activity will occur in
the temporal lobe, such shown in the next figure.
Figure 7.08.
Nerve activity when a "coffee cup" is heard,
as for example being set down upon a table.
The concept "coffee
cup" consists of the simultaneous activation of any, or all, of the three
tiny regions in the three tertiary cortices of the posterior lobes. This
is shown in the next figure.
Figure 7.09.
Nerve activity corresponding to "coffee cup."
The activity pattern
corresponding to "coffee cup" depicted in Fig. 7.09 is said to be "generalized."
That is, there are many specific ways a coffee cup can be perceived, and
indeed there are many variations of coffee cup shape, appearance and sound,
yet they all end up creating the one, generalized pattern "coffee cup."
Frontal Lobes
The brain not only
perceives, it also generates movement. A movement that
is thought about and later commanded is the result of nervous activity in
the frontal lobes. There's a "reverse" pattern for this activity; the process
starts in tertiary cortex, and proceeds in the direction of primary cortex.
This is depicted in the next figure.
Figure 7.10. Flow of nerve activity when some activity is planned and
performed. The flow in this case is from tertiary to primary.
The frontal lobe
architecture is analogous to that of the posterior lobes, in that the most
conceptualized of ideas and plans are created in the frontal tertiary cortex,
which delivers vague "executive" directives to the frontal lobe's secondary
cortex, which formulates more specific action commands and delivers them
(as necessary) to the motor strip. The motor strip requests permission from
the sub‑cortical "reticular activating system" (RAS), and if the RAS approves
the request it is acted upon by sub‑cortical brain areas (Luria, 1973), which
carry out specific actions (orchestrated in detail by the cerebellum).
The frontal lobe's
secondary and tertiary cortices are also referred to by the two terms "prefrontal"
cortex and "pre‑motor" cortex. The prefrontal cortex has undergone the greatest
amount of recent evolution, according to arguments based on the increase
in frontal lobe size versus phylogenetic location (i.e., ratio
of frontal lobe size to total cortex is greatest for humans, next greatest
for chimpanzees, etc). Functions performed by the frontal lobes in humans
are often unique, or most advanced, in humans, whereas most areas in the
posterior lobes have pre‑human analogues. The prefrontal lobes also reveal
their evolutionary recentness by continuing to undergo rewiring until the
age of 5 to 7 (Thatcher, 1997),
and even the "late teens." Giedd and Thompson (2001) write "In late teens,
the prefrontal cortex is the area that's changing the fastest..." (according to neuro-imaging studies). This is consistent
with the general principal that "ontogeny recapitulates phylogeny."
Laterality
The right side of
the body is commanded to move by the left cerebral hemisphere's (frontal lobe)
motor strip. Likewise, the left side of the body is commanded to move by
the right cerebral hemisphere. This left/right crossing‑over architecture
is also adopted by sensory input; sensory information from the right side
maps to the left brain, and visa versa. The reason for this is still a subject
for speculation. The corpus callosum, which interconnects the left and right
cerebral hemispheres, allows for the coordinated movement of both sides of
the body, and also allows for some of the computational results of specialized
areas on one side to be exchanged with related areas on the other side.
Proto‑humans probably
had left/right symmetry, in the sense that the right and left cerebral hemispheres
had identical capabilities, being mirror images of each other in layout. This would have provided redundancy in case one side was
injured (by a fall or blow to the head). Modern humans have asymmetric brains:
the left and right cerebral hemispheres, LB and RB, are somewhat different,
and are "specialized" for certain types of tasks. RB has more long‑distance
inter‑connections than LB, whereas LB has many areas that are highly intra‑connected,
which in turn are connected to other highly intra‑connected regions within
LB.
The best known of
LB's highly intra‑connected areas are Wernicke's Area
(language comprehension) and Broca's Area (language production). Wernicke's
Area is located near the interface of the three
posterior lobes, in LB only (right-most pattern of dots in Fig. 7.11, upper).
Broca's Area is located in the frontal lobe's secondary cortex, in LB only
(left-most pattern of dots in Fig. 7.11, upper). There's a discernible pattern
for the tasks performed in these specialized, highly intra‑connected LB areas: namely, these tasks are inherently sequential, which means
that the temporal order of events is crucial! For example, both receptive
and productive language involves the processing of sequential events (sound
perception and production). Changing word order can profoundly change meaning
("Ed ate the bear" versus "The bear ate Ed."). In contrast, RB tasks are
holistic; they resemble those that a parallel computer processor (neural
network) performs, such as instantaneous image recognition.
Figure 7.11 Upper panel shows location of language comprehension area, Wernicke's
Area (right-most pattern of dots), and speech
production area, Broca's Area (left-most pattern of dots). The lower panel
shows the location of the inferior parietal lobule, IPL, which monitors the
spatial relationship of body parts in relation to the immediate environment.
It is interesting
that RB's counterpart to Wernicke's Area, shown
in Fig. 7.11 (lower panel) is devoted to the task of monitoring the location
of body parts in relation to each other and the immediate physical environment.
This area, called the "inferior parietal lobule," or IPL, plays a critical
role during manual interactions with the environment, such as reaching out
to pick fruit from a nearby branch.
It is tempting to
conjecture that before humans were capable of speech the left hemisphere's
IPL counterpart region also functioned like the present‑day IPL in RB. Because
reaching out to pick fruit had sequential components, it would have been
natural for mutations to modify what once was an LB IPL in a way that later
presented an opportunity for further modification that led to a simple form
of language capability. This region must have been built-upon to produce our
present‑day Wernicke's Area, which plays a critical
role in language comprehension. This task consists of monitoring the relationship
of sound percepts to each other over time, somewhat similar to the way the
RB's IPL monitors body part relationships over time. As Wernicke's Area evolved in LB, it must have gradually displaced
the former IPL function.
A great deal of
public interest was generated during the 1970s and 1980s by reports of LB
and RB differences, or lateralization. For example, RB is described as being
intuitive, holistic, inductive, timeless, visuo‑spatial, non‑verbal and pessimistic,
whereas LB is described as being verbal, analytic, logical, rational, time‑oriented,
deductive and optimistic.
Traditional psychologists
must have resented the newcomers to their field who used instruments to measure
things, and who used rigorous techniques to study long-standing matters
that had been the subject of arm chair speculation. The old-fashioned psychologists
accused those who studied split brain patients, and found LB and RB differences,
as suffering from “dichotomania” – as if the new investigators were over-interpreting
their data due to an excess of enthusiasm. But the data is convincing, and
often dramatic.
When LB is damaged
(or when it is temporarily disabled by sodium pentathol injected into the
left carotid artery) the patient's speech capability is almost non‑existent.
Curiously, though, the still‑functioning RB does what it's able to do speechwise:
the patient can swear, utter emotion‑laden pat phrases, sing songs with the
right words, and recite the alphabet. RB cannot (usually) put together a
sentence, since grammar capability resides in LB.
Occasionally, a
patient whose corpus callosum has been cut can still manage to communicate
in a simple way using the rudiments of grammar. These cases offer very interesting
insights into the differing "personalities" of LB and RB. One famous example
was reported by Gazziniga (1978) which suggests that LB and RB can have different
goals in life. Their oft‑used subject P.S. was questioned about his job
choice in an experiment that allowed only RB to answer, and "automobile race"
was spelled out. As Gazziniga writes "This is most interesting,
because the left hemisphere frequently asserts that he wants to be a draftsman"
(p. 143). How poignant!
Chicken Claw Experiment
My favorite
illustration of the independent operations of LB and RB has been referred
to as “the chicken claw experiment.” This experiment was conducted by Michael
S. Gazzaniga and Joseph E. LeDoux using patient P. S., who had undergone
a full callosal surgery (cutting of the corpus callosum, interconnecting LB
and RB) to control seizures. I shall quote from descriptions appearing in
two books: Gazzaniga and LeDoux (1978) and Gazzaniga (1985).
Two problems are
presented simultaneously, one to the talking left brain and one to the non-talking
right brain. The answers for each problem are available in full view in front
of the patient. Gazzaniga and LeDoux
(1978).
Figure
7.12. “Chicken claw experiment.” The “task” (top) has two parts,
presented to a brain half. The answer choices, below, are in full view to
both brain halves.
...the experiment
requires each hemisphere to solve a simple conceptual problem. A distinct
picture is lateralized to one hemisphere: in this case, the left sees a picture
of a claw. At the same time the right hemisphere sees a picture of a snow
scene. Placed in front of the patient are a series of cards that serve as
possible answers to the implicit questions of what goes with what. The correct
answer for the left hemisphere is a chicken. The answer for the right hemisphere
is a shovel.
After the
two pictures are flashed to each half-brain, the subjects are required to
point to the answers. A typical response is that of P.S., who pointed to
the chicken with his right hand [controlled by the left brain] and the
shovel with the left [controlled by the right brain]. After
his response we asked him, "Paul, why did you do that?" Paul looked up, and
without a moment's hesitation said from his left hemisphere, "Oh, that's easy. The
chicken claw goes with the chicken and you need a shovel to clean out the
chicken coop."
It is hard
to describe the spell-binding power of seeing such things.
My interpretation
is that the normal brain is organized into modular-processing systems, hundreds
of them or maybe thousands, and that these modules can usually express themselves
only through real action, not through verbal communication. Gazzaniga (1985).
... a basic mental mechanism common to us all. We feel that
the conscious verbal self is not always privy to the origin of our actions,
and when it observes the person behaving for unknown reasons, it attributes
causes to the action as if it knows, but in fact it does not. It is as if
the verbal self looks out and sees what the person is doing, and from that
knowledge it interprets a reality. Gazzaniga and LeDoux
(1978).
Frontal Lobes
The frontal lobes
play a key role in orchestrating behaviors associated with LB/RB specializations.
For example, RB prefrontal (RBF) originates emotional outbursts, whereas
LB prefrontal (LBF) works to produce socially responsible behavior. The limbic
system appears to be more strongly connected to RBF, and uses it to elaborate
emotionally driven behaviors. LBF, on the other hand, appears to be the seat
of the "conscience" and inhibits any RBF desires for socially inappropriate
behaviors.
This was dramatically
illustrated by the famous case of Phineas Gage, who suffered a railway construction
accident in 1848 that caused a metal tamping rod to explosively penetrate
and destroy his LBF (and a small part of RBF). Without the inhibiting effect
of LBF upon RBF, his behavior was "fitful, irreverent, indulging at times
in the grossest profanity... at times pertinaciously obstinate... he has
the animal passions of a strong man." (Harlow, 1868). This old example
illustrates the well known finding that RB's language ability is usually
limited to profanity, songs and other memorized verbal material, such as the
alphabet. A wealth of studies show that LBF is the site of the most advanced
and human traits, such as conscientiousness, positive social behavior, rationality,
strategic planning, and positive affect (mood). LBF is often referred to
as the site of "executive function." RBF, by contrast, is associated with
lack of inhibition, anti‑social behavior, emotionality, and negative affect.
RBF is more closely connected to the sub‑cortical limbic system, the source
of emotions.
If RBF and LBF could
take positions concerning the idea that "the genes enslave us for their sometimes
pernicious activities, and that individuals should rise up and become liberated
from this genetic enslavement," it is obvious which side LBF and RBF would
be on, and they wouldn't be on the same side! More on
this in a later chapter.
This chapter's brief
description of cerebral architecture, and the functional relationships of
components, is part of the accepted neuropsychology literature. Every normal
person's brain functions this way. If the brain was a "blank slate," as Francis
Bacon initially suggested, and philosopher John Locke systematically expounded,
then how amazing it would be for the blank slate to form itself into the
same well‑defined areas, with corresponding functions, in all people ‑ regardless
of their individual upbringing and environmental experiences! This old idea
is best forgotten. Even Bacon and Locke would probably disown the outdated
notion if they were alive today and could know about recent neuropsychology
findings.
The genes assemble
brains with the same architecture, modules and functional relationships,
and this process occurs automatically ‑ shall I say "mechanistically." This
view of the brain is consistent with the reductionist theme found throughout
this book. The next chapter is more speculative as it treats the brain’s role
in evolution.

─────────────────────────────────
CHAPTER
8
─────────────────────────────────
THE BRAIN'S ROLE
IN EVOLUTION
"Aristotle was famous
for knowing everything. He taught that the brain exists merely to cool the
blood and is not involved in the process of thinking. This is true only of
certain people." Will Cuffy.
The brain is assembled
by many genes. Each gene has had to establish itself within a species genome
that, by definition, was successful at the time the new gene competed for
a place in the gene pool. We should assume that each brain‑affecting gene
established itself in the human genome at a different time from all other
brain‑affecting genes. Obviously, all genes achieve their success without
the benefit of how well it might work with any future gene. Each successful
gene has had to compete with existing genes, or at least provide a benefit
that exceeds penalties from incompatibilities with existing genes. From the
perspective of the gene, the individual's brain has the responsibility of
spreading the gene widely into future generations. This is another way to
express the unavoidable tautological assessment that a gene's job is to
try to infiltrate the species genome and persist forever.
When a new gene
modifies the hardwired neural connections of some brain region (by creating
new connections between neurons or by changing the size of synapses of existing
connections), the function of the modified brain region is likely to be in
conflict with other brain regions. Since the purpose of the brain is to
influence behavior on behalf of the genes, brain regions necessarily are
in competition with other brain regions for influencing behavior. Rarely
is the individual aware of this conflict. When the conflict is extreme,
when it affects emotional state, we might say that the brain is in an unsettled
state of "cognitive dissonance." Almost all competitions for influencing
thought and behavior are worked out peacefully below conscious awareness.
The Brain as a Mechanism
The brain is a mechanism,
albeit a "wet chemistry" mechanism. Just as all chemical interactions are
merely physical interactions at the atomic and molecular level, so are all
brain interactions ultimately the working out of physical relationships between
atoms and molecules. When we say that current flows along a neuron's axon,
we refer to a physical process of the axon's membrane becoming more permeable
to sodium atoms, allowing charged atoms to enter the axon from the surrounding
fluid, etc. Every motion of every atom is governed by a =
F/m and quantum physics (as explained in Chapter 1). It would be cumbersome to try to understand brain function
by invoking this basic level of physics since such a task would be incomprehensibly
difficult. Wet chemistry is a less cumbersome level, but still too daunting
for most brain studies. A more tractable, and hence powerful, level for understanding
brain function is to think in terms of neural networks.
A neural network
is a partially interconnected group of neurons. One
network may also have connections with other neural networks. The term "partially
interconnected" is important, for it is the genes that determine the overall
pattern of which connections exist. A "fully connected" network is impractical
when the number of elements (neurons) exceeds a few hundred, since the number
of possible connections between elements grows as "N‑1 factorial." Synaptic
connections between neurons are either excitatory or inhibitory. In the brain
a neuron may have many synaptic connections to a specific target neuron,
and absolutely no direct connections to most other neurons.
Consider all neurons
in the network that are connected to one individual neuron. At any moment
some of them will be in the process of "discharging," causing their synaptic
connections to other neurons to become active (releasing neurotransmitters
across a synaptic gap). Each target neuron sums the excitatory and inhibitory
discharges on its cell body, and if this sum exceeds a threshold it in turn
discharges, causing neighboring neurons with which it has output connections
to possibly also discharge by the same process that led to its discharge.
A neural network can be made to "resonate," which is a way of stating that
a pattern of firings within the network continues for many clock cycles (tens
of milliseconds in the brain) once triggered by an appropriate stimulation
from the connections that the neural network has with neighboring neurons
(or neural sub‑networks). All of this is well understood by neural network
specialists, and I provided a brief introduction of it here to give the reader
a taste for the mechanistic, or reductionist nature of brain phenomena.
An even more useful
level for understanding brain function is to speak of brain regions in terms
of their function. When we use such terms as "the reticular activating system"
(RAS) we know that the elaborate neural network explanation for the region's
function is theoretically possible but at the present state of brain understanding
these a = F/m
ways of accounting for a regions function are not very feasible or even useful.
So we proceed by saying, with blatant anthropomorphism, that a cortical region
sends a "request for activation" to the RAS, and if RAS "grants the request"
this originating cortical area becomes more active, and this activity enables
it to increases it's inhibition of "competing" cortical brain areas, allowing
it to succeed in "achieving behavioral expression." Even that way of speaking
is cumbersome, but it captures the flavor of the mechanistic competition
of one cortical neural network, having gene‑directed hard wirings, with a
neighboring cortical neural network, having other gene‑directed hard wirings.
In any description
that attempts to achieve brevity, such as this one, there are many unmentioned
details about which a specialist could complain when they are left out. Sure,
I didn't mention neurotransmitters, and their re‑uptake, or their breakdown,
and dozens of other things going on, but they are all mere elaborations of
the same basic physical mechanism. Additional details are too numerous to
mention, but also too similar in terms of their ultimately physical action
to warrant mention for present purposes.
I have risked boring
you with some physics of the brain in order to show how in principle brain
function can be understood as coming under the influence of the genes. For it is the genes that direct
the process of "pre‑wiring" the brain. Initially, too many connections are
created, and for several years after birth approximately half of the neurons
and their connections wither and are lost. But the starting point at approximately
birth and some years later (depending on the brain region), the overall placement
of neuron type and the majority of connections from each neuron to others,
is supervised in a general way by the genes. Some genes influence one region
(i.e., a neural network) and not others, while other genes influence several
different specific regions. Any single neural network is most likely the
result of several genes.
This way of viewing
brain development, emphasizing as it does the role of evolutionary forces
on the architecture and interconnectedness of the brain, leads to a perspective
in which overall brain function is the working out of a competition of mental
modules, each endeavoring to express itself by
maximizing its influence over behavior. The "modularity of mentality" perspective,
in which modules compete with certain others, is still controversial (for
reasons I don't understand). And the idea of connecting specific modules
to specific genes is so amorphous a speculation that it is not yet a sub‑discipline
of the brain sciences. Evolutionary psychologists adopt this view (see
Barkow et al, 1992), and it seems inevitable to me that
sometime in the 21st century neuropsychologists will also, and maybe late
in the 3rd millennium people who call themselves psychologists will come
aboard.
Recent Evolutionary
Hotspots in the Human Brain
In humans the prefrontal
cortex is proportionately larger than the rest of the brain compared with
all other animals. Thus, there's an evolutionary trend revealing that the
prefrontal cortex has been the focus of recent human evolutionary adaptations.
This makes the prefrontal cortex one of the most interesting brain areas
to understand.
Comparison with
other mammals reveals that the tertiary cortices of the posterior lobes are
also proportionately larger in humans, indicating that they also have been
undergoing rapid evolution in recent evolutionary time. The most obvious
example is Wernicke's Area, located in the temporal
lobe's tertiary cortex. So add LB posterior lobe tertiary cortical areas to
the list of interesting human evolutionary "hotspots."
Are there any evolutionary
hotspots in the human right brain posterior lobes? The short answer is "no."
It therefore seems that the left brain has evolved more during human history
than the right. It is even tempting to suggest that what distinguishes humans
from other animals is their left brain.
Note one qualification
that applies to most usages of the terms "left brain" and "right brain": about 2% of the population has laterality reversed. In
these people language and other sequential tasks are performed by areas in
their right brain, and holistic functions are performed by their left brains.
Most of these people are left‑handed with the unhooked writing position.
Neuropsychologists use the terms right brain and left brain to refer to the
specializations found in that 98% of the population with "normal" lateralization.
So, whenever the terms LB and RB are used, think of the left and right brains
of the 98% of people who possess the normal lateralization.
Why is the Left
Brain Evolving Faster?
What is it about
the left brain that gave it the greater burden for advancing human evolution?
One clue comes from the microscope. The left brain isn’t as "white" because
fewer neurons are coated with an electrical insulator composed of a whitish,
fatty substance called myelin. The greater myelinization of the right side
is required by the greater proportion of right side neurons that connect
with distant neurons through long axons. In contrast, neurons on the left
side are more often connected to nearby neurons, and therefore require less
insulating myelin.
But what does this
mean? The left brain is characterized by a neural architecture in which isolated
neural networks perform their specialized tasks and then communicate their
results among themselves through a smaller network of interconnections. Functionally,
this is a better architecture for performing sequential tasks. Language
is a good example. A sentence consists of a sequence of sounds that have
to be in their proper place in order to convey the intended meaning.
Some have speculated
that the evolution of lateralization started with our fruit eating ancestors,
who would use their left arm and hand to support themselves and maintain
balance while the right hand reached out to pick fruit. Fruit picking is somewhat
sequential, as the hand must be guided by the eyes to reach for the ripe‑colored
fruit, grasp it with fingers using just the right force, tear it off the
branch, and then bring it to the mouth for eating. Recall that the sequentially
performing right hand is controlled by the left brain, which would therefore
be the one requiring a sequential neuronal architecture.
If embryological
development provided for a sequential brain architecture in one part of LB
because it evolved in that location by chance for the purpose of picking
fruit, then when another sequential task became adaptive the forces of evolution
would more often find a favorable mutation of genes that code for the left
brain, since fewer mutations would be needed to add to a pre‑existing architectural
capability for the task of producing a new sequential capability. This, according
to one speculation, is why the left brain took on most new sequential tasks
presented to it by subsequent evolutionary opportunities.
Brain "Dominance"
Damaging brain strokes
in LB tend to produce more noticeable deficits than those in RB. This is
because LB performs language tasks. For this reason, unfortunately, it has
become customary to regard LB as the "dominant" hemisphere. But, to call
LB dominant over RB for this superficial reason is misleading!
The limbic system
(that drives emotions) influences the RB frontal lobe more strongly than
the LB frontal lobe. This makes RB a better candidate for playing a dominant
role. RB gives overall shape to behavior, while LB is relegated to a supporting
role. When LB began its sequential specializations it must have been a useful
"tool" for RB (which in turn was a tool for the limbic system, which in turn
was a tool for the genes). "Values" are more likely to originate with RB,
and I claim that the genes have put in place more of their “agenda protection
circuits” in RB. The natural condition, I suggest, is for RB to be "in
control," using LB to help achieve genetic ends. This important thought
will come up repeatedly in subsequent chapters, and it is a basis for the
individual to design strategies for individual liberation from the genes!
Even though RB has
control over decisions that matter to the genes, I believe the seat of consciousness
is in LB's tertiary cortex. This may seem to be a curious arrangement, but
upon further thought it makes sense. We associate consciousness with planning
future activities. Recall that brain structures with a more ancient origin
usually have veto power over behaviors, as when RAS handles requests for
action and either "authorizes" or "vetos" them. It may happen that RB tasks
LB with imagining future scenarios and their likely consequences, while RB,
in close consultation with the (emotional) limbic system, then makes a “judgment
call” and decides whether or not to proceed with the plan of action that
was under consideration. After imagining scenarios, one may be accepted while
others are vetoed; all of this may occur at a subconscious level, with RB
working in conjunction with the ultimate authority: the limbic system. LB
must make sense of the outcome, so it "confabulates"
an explanation for the chosen plan of action. Michael Gazzaniga names the
left prefrontal module that performs this confabulation the "interpreter"
(see Gazzaniga, 1978, p. 146; especially Gazzaniga, 1988, p. 229; and Gazzaniga,
1992, p. 121).
Rational thought
has become an ever more important tool for evaluating the consequences of
hypothetical actions. This is why LB must have been such a "hot‑spot" for
human evolution for probably the past 130,000 years, and especially the past
12,000 years.
Before LB began
to evolve its unique specializations, perhaps 250,000 years ago, the function
performed by a damaged area of one side could be easily assumed by the counterpart
area of the other side (relying upon the corpus callosum for inter-hemispheric
communication). Lateralization brought with it risks of lost redundancy,
yet this loss was apparently smaller than the gains from being able to solve
problems that were common in the late Pleistocene and Holocene. We must assume
that some important need started the selection for LB specializations. It
may have been the payoffs for improved tool‑making, language, or dealing with
a more complicated social setting that required logical thinking skills (such
as "theory of mind" abilities).
Whatever the original
impetus for LB specialization, it seems to have assumed the new duties as
if forsaking redundancy with abandon. Just consider the list of important
LB skills that are unique to humans: verbal, analytic,
logical, rational, time‑oriented and deductive skills. It seems inescapable
that LB has acquired more recently‑evolved, distinctly human adaptations
than RB. When you damage LB (posterior lobes), you get a regressed, more primitive
person; but when you damage RB (posterior lobes), you get someone handicapped
in mostly long‑standing, primitive traits.
Since recently-evolved
traits are the least entrenched, and are most subject to disruption by the
latest mutations, we should expect to encounter a wider variation of ability
for the recently evolved traits than long‑established ones. This view correctly
predicts that literacy, being a recent human achievement, should be more
variable than other abilities; whereas verbal language ability, having started
its evolution much earlier, should be more robust. It also explains why everyone
is capable of anger, fear, sexual arousal, and jealousy, while some people
are deficient in logical, rational and analytic ability.
Brain Modules and
Genes
As with any organ,
no single gene codes for the construction and function of an entire organ;
many genes contribute. When several genes contribute to the same trait, they
constitute a group of "polygenes." For example, one gene may play a major
role in forming the heart's left ventricle, with minor support from other
genes; another gene may have major responsibility for assemble of the right
ventricle, but also contribute to the left ventricle's assembly. Both genes
belong to a polygene group for constructing the heart.
The same argument
applies to the brain. Many genes are required to assemble the primitive brain
stem's reticular activating system, for example. Others assemble various
parts of the limbic system. Finally, other genes assemble the surrounding
neo‑cortex, LB and RB, and the interconnecting corpus callosum.
All brain components
are interconnected with other components, and they function together as if
they were "designed" to work together. When the various components work together
it is because they have been present in the genome together long enough
to adapt to each other's presence. Initially, when a new brain component
is mutating into existence, it is useful to understand that the pre‑existing
components were not meant to work together with the new component. Each
new "addition" occurs against a background of pre‑existing brain components
which had worked together successfully prior to the appearance of the new
component. As components appear, they, as well
as the pre‑existing components, co‑evolve to enhance the working relationship.
When our ancestors
began to lose their fur, the fur altering allele had to co‑evolve with the
gene(s) that made furry babies irresistibly attractive to mothers. There
are many baby features that cue the mother to act like a mother, and the lack
of fur amidst all the other baby features must have been disconcerting to
mothers during the transition. Today, a cat resembles a primitive baby in
size, weight and furriness. The fact that many people find cats irresistible,
and sometimes hold them like a baby and speak "motherese" to them, suggests
that the ancient collection of cues for eliciting mothering behaviors still
exist in some residual form.
As the gene for
a new brain module is selected, it evolves to be compatible with pre‑existing
modules, and the genes for the pre‑existing modules simultaneously undergo
modification in response to the new module. The evolution of genes that affect
the brain is governed by the consequences each gene allele has on the success
of the individuals carrying the genes to survive and reproduce. Or, to be
more rigorous, a brain‑related gene allele's success depends on its ability
to produce phenotypic changes that work with the prevailing phenotype in
a way that enhances the individual's success in delivering all of its genes
to future generations, under a typical range of environments.
Since there are
many ways to construct any organ, there will be many potential competitions
between genes. An allele that produces a larger heart ventricle is in competition
with alleles that produce smaller ventricles. Natural selection achieves
a better heart by rewarding individuals having the better heart, and thereby
rewarding those gene alleles responsible for producing the "better" ventricle
size.
Whereas it may be
easy to comprehend how a gene that codes for anatomy, such as heart ventricle
size, can be in competition with another gene, it is more difficult to imagine
the competition between genes that assemble brain circuits governing behavior;
nevertheless, it happens. A brain gene may be in competition with another
allele, even while it is "cooperating" with a different set of brain genes.
(Excuse the anthropomorphizing; if it bothers you just convert my brief descriptions
to a rigorous lengthy one).
Genes compete for
phenotypic expression at impressively high conceptual levels. Language ability
evolved by creating proto‑Wernicke's Area circuits and proto‑Broca's Area
circuits within LB (plus other cortical areas, interconnections and anatomy
modifications). This was a major accomplishment, involving many small incremental
steps. Other frontal lobe traits, such as assertiveness, aggressiveness,
nurturance, empathy and altruism, are under significant genetic control, accounting
for approximately 50% of observed variance (Rushton, 1997).
It has once again
become fashionable to think of brain function as being "localized." Although
"phrenology" deserved to be discredited, it's
ultimate theme was correct: namely, that most attributes of brain function
are determined by activity in specific brain regions. They were wrong to place
"combativeness" where the temporal and occipital lobes join, for example,
but it is localized, and belongs in the prefrontal lobes (probably in RB).
Many functions require the participation of several specific areas. Productive
language is a well‑studied example, exhibiting involvement of specific parts
of the left frontal, temporal, and parietal lobes. Physical damage to each
region produces specific, predictable language deficits. This means that something
as complicated as language requires the cooperation of regions with specialized
capabilities, and the fact that these regions aren't next to each other,
but are located in different lobes, does not undermine the view that brain
functions are localized.
This reductionist
way of viewing brain function is supported by the notion that a finite number
of genes assemble the brain. Polygenes create brain modules, consisting of
specific physical networks of interconnected neurons along with an approximate
set of synapse sizes.
Mental function,
like brain architecture, appears to be modular (Fodor, 1983, Gardner, 1983,
Gazzaniga, 1985, Restak, 1986, Cosmides, 1989, Cosmides and Tooby, 1992,
Restak, 1994). Granted, the modules interact with each other, but they can
be usefully considered as modules with functional specifications. Consider
the analogy of a system's analyst parceling out the task of writing a large
computer program to several teams of programmers. Each team is charged with
delivering a module of code that meets functional specifications. A programming
team is like a gene, their code is like a hard‑wired brain module, the function
performed by the module is like a mental module, the joining of modules is
performed by the systems analyst, and the running of the completed program
code is like a brain performing mental tasks.
Modules Compete
With Each Other
Modules compete
with each other for "expression." For example, one area in the occipital
lobe may be able to correctly perceive and identify an object from its visual
appearance, a cup for example, while another area in the parietal lobe may
be able to identify the same object by its tactile feel, and another area
in the temporal lobe may be able to infer the same object's identity from
the sound it makes when set down upon a table top. Each will produce a signal
of recognition when the necessary stimuli are presented, and somewhere in
the adjacent tertiary cortical regions (where the three posterior lobes merge)
the object identification of "coffee cup" is made.
It would be ridiculous
to conclude from this capability that there's a "coffee cup recognition"
gene. Rather, there's a polygene‑created module for recognizing curved shapes,
another for shadings that contain surface topography information, etc. These modules are interconnected so that experience with
the real world, or at least one that contains coffee cups, allows synapse
strengths to be modified such that when a coffee cup with arbitrary orientation
is viewed the various percepts are joined together to trigger the perception
"coffee cup."
The brain's experience
with the real world adjusts synapse strengths so that no other region will
be triggered to "resonant" activity when a cup is presented to the posterior
lobes' primary cortex (sensory input) areas. If a totally unfamiliar object
is presented, there will be a competition to identify it. When shown a German
beer stein for the first time, the occipital lobe (sight) may report "something
like a vase," the parietal lobe (feeling) may report "something like a
large‑handled cup" and the temporal lobe (hearing) might report "like a
brick." The discrepant reports would compete, as the frontal lobe might
want to engage in further exploration to resolve the discrepancy (which
is a job for “consciousness).

Figure 8.01. Reversible Goblet, illustrating competition between brain
regions vying for "acceptance" of their respective interpretations.
Look for the two dark face profiles, facing each other.
The many examples
of images with "figure/ground reversal" conveys in the most dramatic way
how competing modules strive to prevail in having their "interpretation" accepted.
Escher drawings (Escher, 1961) exhibit a wealth of figure/ground perceptual
competition.
Identifying situations,
such as a "social situation," is subject to the same perceptual competition,
although the frontal lobes will play a more active role in generating competing
hypotheses. Context may be an important "input." "Do I know the person? Does he have a hostile stance? Does
he have comrades?"
Consider the metaphor
of a school classroom for understanding brain module competitions. The teacher
poses a question, and the students try to understand the question and come
up with an answer. Some students will both understand the question and have
a possible answer, and they will raise their hand. The teacher calls upon
a student to present an answer, quite often it's the student whose hand is
waving most excitedly (or maybe the student with the best past performance),
and after hearing the answer she passes judgment. If it is incorrect or inadequate,
the teacher calls upon another student.
This classroom example
is a good metaphor for how the brain works. When a person is presented with
an unusual situation, some modules in the brain "recognize" something, and
they request activation by the RAS (reticular activating system). The RAS,
working in coordination with a higher level cortical system that keeps score
of previous successes and failures, tentatively authorizes a module to "present
its case" for evaluation. The module that wins the first round for presenting
its interpretation may be the one that most strongly felt it understood the
situation and had the correct interpretation (like the student who waved
his hand most excitedly); its request to RAS may have been the strongest
among the competing modules. When the first module presents its interpretation,
some type of evaluation occurs (perhaps involving the reaction of other modules),
and this interpretation may be accepted, or it may be tentatively rejected.
If it is rejected, or set aside, another round of RAS requests for activation
is performed, and another judgment is made. At some point, a winner is declared,
and the winning module's interpretation is what serves as the basis for any
required action. The losing modules do not simply stop trying to compete
for RAS attention, however. As more perceptions occur,
or as behaviors either validate the accepted interpretation or invalidate
it, the other modules are ever‑ready to renew their claim for being heard.
The example of the "reversible goblet" shows how this process "feels" for
the domain of visual interpretations.
The process of generating
behavior is also a competition between competing frontal lobe modules. "Shall
I turn and run? Or scream? Or attack?" Imagine that one person may inherit
a propensity to "attack" in ambiguous social encounters, while another may
be genetically inclined to "run away." Just as animals have inborn temperaments,
so do humans. And the mental process that precedes an action consists of
a competition between brain areas. To the extent that one brain area is
assembled by a different polygene group than another brain area, which is
inclined to a different type of behavioral response, the genes primarily
responsible for wiring the competing brain modules are competing with each
other for behavioral expression. The same classroom metaphor described
above can be used to understand this situation. After a situation is understood,
and when action is necessary, the frontal lobe modules will compete for expression
(i.e., control of behavior) in the same manner that the
"understanding and interpretation of the situation" modules competed. The
RAS (another part of RAS than used for “adjudicating” perceptions) receives
requests for action, and eventually one module's proposed action is "accepted"
(given “authorization” for initiating a behavior).
Thus, both perceptions
and behaviors exhibit the quality of involving several mental modules in
competition for acceptance and expression.
Does it matter whether
the brain accepts, and acts upon, the perception that the sky is angry and
the wind god and sun god are arguing, versus the competing perception that
the wind is bringing clouds from somewhere which cover the sun and may cause
rain? In the contemporary world it can matter more than it did in the ancestral
world. For this reason, it matters whether RB interpretations versus LB interpretations
tend to gain acceptance in an individual's brain. To the extent that genes
wire brains to be predisposed to some "interpretation styles" over others,
the respective genes are in competition. The next chapter will deal with
this subject in greater detail.
Intelligence and
IQ
“IQ is what IQ tests
measure!” It should be emphasized that IQ, as measured this way, is just
one of many components of what most people refer to by the term "intelligence."
Ironically, IQ is not a prefrontal function. Prefrontal lesions do not reduce
IQ; indeed, in some cases frontal lesions have enhanced IQ. This enhanced
performance could be explained by a theory that views the frontal lobes as
being prone to "interfere" with posterior lobe performance (such as a tennis
player "thinking" too much); by injuring a prefrontal lobe the posterior
lobes are freer to perform unhindered, boosting measured IQ. The WAIS (Wechsler
Adult Intelligence Scale) IQ test has two parts: the "verbal" part and
"performance" part, and these parts probe left and right (posterior) brain
function, respectively. The Woodcock‑Johnson has two parts, also probing
left and right (posterior) lobes. The WAIS verbal and performance IQ scores
differ by 3 points, on average. A difference of 10 points should occur in
only 5% of cases; and differences larger than this are usually caused by
a lesion to one side of a posterior lobe.
This concordance
of IQ scores that separately probe LB- and RB‑function invite speculation
on the number of genes that affect posterior lobe capability on both sides.
However, it is possible that a small number of genes contribute to "general"
intelligence, and the rest contribute to specific abilities. This is consistent
with the finding that a person's profile of subtests will have a pattern,
with some parts of the "verbal" being low, and others high, while the average
of the verbal parts average about the same as the average of the performance
subtest parts. Psychometricians continue to find it useful to make a distinction
between specific sub‑test performances and Spearman's (1927) "g factor" of
mental ability. Tests that identify g‑loaded performance afford better correlations
with genetic relatedness (i.e., identical versus paternal
twins), and g‑loaded test scores are better predictors of academic performance
than standard IQ tests.
Tests have been
developed for assessing frontal lobe performance. The Halstead‑ Reitan Battery
includes tests of frontal lobe assessment. Components of the Montreal Neurological
Institute Battery, and also Luria's Neuropsychological Investigation, also
test for frontal lobe function. The frontal lobes are so complex that no
single test can capture all significant features. For example, effective business
executives have especially capable frontal lobes, and they excel in the development,
evaluation and implementation of "big picture" strategies. The business "world
of hard knocks" reveals who some of these especially well‑endowed frontal
lobe "executive function" people are. Bill Gates, Steven Spielberg and Lee
Iacoca are examples. It would be interesting to know if they would have been
identified in childhood as having especially talented frontal lobes using
existing tests purporting to probe frontal lobe function. Some day, tests
for executive function may capture this elusive capability.
Whereas someone
like Bill Gates must have superior scores for both frontal and posterior
lobe function, it must occasionally occur that people are born with disparities.
For example, president Jack Kennedy is supposed
to have scored a mere 125 on IQ tests. He obviously would have scored higher
on any executive frontal lobe test. It may be more common for people to be
born with the opposite disparity, in which posterior lobe IQ is higher than
frontal lobe executive ability. Indeed, this could be the more common disparity
because frontal lobe function is a more recent focus in human evolution.
The ability to create culture, and to absorb and use cultural elements that
other people are observed to use, must have been an important pressure for
human evolutionary selection during the past 60,000 years. This idea will
be taken up in a later chapter.
Number of Brain
Genes
It has been estimated
that as many as half of the entire set of human genes have some influence
upon intelligence (Weschler, 1974, as cited by Seligman, 1992). For the calculations
that follow, I will assume that 30% of human genes affect the brain. In
theory, every aspect of brain function can be associated with a gene that
has the most control over it. (To call such a gene the "whatever trait gene"
overlooks another fact, that the same gene probably affects several other
phenotypic traits which are sometimes unrelated to the main trait; this
is referred to as "pleiotropy.")
If the human genome
consists of 22,000 genes (functional sections in the “junk DNA” regions will
also be present), and if humans share 98.77% of genes with living chimpanzees,
then humans differ from chimpanzees at approximately 270 gene locations (1.23%
of 22,000). Of these 270 genes, probably more than 30% have some influence
over the brain's development. Let us assume that 80 genes are responsible
for making the human brain different from the chimpanzee brain.
If the common ancestor
for modern humans lived 200,000 years ago, and if the human/chimpanzee evolutionary
split occurred 6 million years ago, then it is possible to estimate the number
of brain genes that are more recent than 200,000 years to be 80*0.2/6 =
2.7 genes. This absurd result requires a few caveats. First, this calculation
assumes that the pace of evolutionary change has been constant during the
past 6 million years. Human evolution may have proceeded faster during the
past 200,000 years than before this time, and the brain is likely to have
been the focus of more than 30% of this evolutionary change, considering
that major human brain expansions occurred at about 1.8 and 0.5 million years
ago (Aielo and Dunbar, 1993). But the most important qualifier of this argument
derives from the restrictive definition of a “gene.” A gene is defined as
a stretch of DNA that “contains the instructions for the production of a
particular protein.” Recent research has shown that non-gene DNA (in the areas
once referred to as “junk DNA”) can affect the expression of genes. Not enough
is known about the number of these locations to include them in the present
argument, so allow me to proceed while keeping this one important caveat
in mind.
If during the past
200,000 years 80% of the genes that were actively evolving were brain‑related,
then the 2.7 multi‑allelic number increases to 7. If the evolutionary pace
for recent times (the past 200,000 years) versus before (6 million years
to 200,000 years) is greater by the factor of 10, then there could be 70 multi‑allelic
gene sites that affect the brain. This number is compatible with the estimate
that humans and chimpanzees differ at 270 gene sites. Since each site may
have many more than 2 alleles per locus, there could be 100 to 200 alleles
whose main effect is on the brain and which are still vying for a presence
in the human genome. This may seem like a small number of gene sites, but
there are 2N combinations of configurations when each site has
two possible states, and if N = 100, there are 1030 such states.
That's an incredibly large number, being larger than the human population
by the factor 1020 (a one followed by 20 zeros)!
The point of these
calculations is to prepare the case for stating that perhaps half of the
present human genetic diversity, and genetic competition (among perhaps several
hundred alleles), pertains mainly to the brain. The brain is a major focus
for ongoing evolution for Homo sapiens.
Recent Brain Evolution
Two lines of evidence
point to the LB‑frontal region as being the site for most of the recent human
evolutionary activity (when I use the term "frontal" I usually mean "prefrontal"
‑ which is common usage). First, this cortical region is where executive
function resides, and executive function is an especially well‑developed,
some would say unique, human capability. Second, the human individual's ontogenetic
development's last big push is in the prefrontal brain areas; and to the
extent that individual ontogeny recapitulates species phylogeny, our most
recent ancestors must have been busily improving the prefrontal cortices,
with increases in both size and function.
RB‑frontal and LB‑frontal
are probably the most recently evolved regions, and I am suggesting that
they gradually came to be in almost constant competition. LB‑frontal has
evolved to supercede RB‑frontal for many tasks (language replacing gestures
being the most prominent). LB‑frontal is capable of inhibiting RB‑frontal
when an appropriate occasion arises. But there are dangers in giving a new
tool too much power, especially when it is strategically positioned to formulate
near‑term strategies as well as long‑term life goals ‑ the way LB‑frontal
is. The genes will find RB‑frontal a more useful agent for controlling LB‑frontal's
"new ideas." Presently, RB‑frontal is probably charged with preventing LB‑frontal
from thinking thoughts that threaten the genetic agenda. Without understanding
any of this, RB‑frontal has taken on the role of acting as an agent for
the genes, and LB‑frontal has unknowingly stumbled into the position of having
the capability for acting as an agent for the individual wishing to liberate
himself from genetic tasks!
It is common knowledge
that people tend to think with certain styles, such that if you know one
of their beliefs or interests you can predict others. Dichotomies abound: Spiritualism versus Rationality, Religion versus Science,
Conservatism versus Liberalism. How curious that
we can expect almost universal agreement in making these aggregations. Consider
the following joke: A prize will be given to anyone who is able
to survive driving from Houston
to El Paso
in a Volkswagen minibus with stickers that read “Down with the NRA” and “make
peace, not war” and “vegetarians rule” and “Vote Democratic.” (I maintain that
it’s possible to predict a person's political party affiliation by knowing
whether they deal with weeds by pulling them or spraying them with weed killer.)
The apparent pattern
of two ways of thinking corresponds to the preferred styles of RB and LB,
and it is obvious which trait corresponds to which brain half. The person
who exhibits RB styles can be thought of as belonging to a group of people
who are "opposed" to the group of people exhibiting LB styles. Just as there
is a competition between an old RB and a new LB within each individual, so
is there a "competition" for prevailing over a society's culture between
RB‑style people and LB‑style people. The genes that wire‑up
RB‑styled and LB‑styled individuals are, as groups, in competition with each
other. C.P. Snow wrote The Two Cultures and the Scientific Revolution
(1961) to call attention to a conflict between two types of people, the literary
academic and the scientist. The book was based on decades of interacting
with both types, and it was written before the great burst of neuropsychological
insights of the 1960s, 1970s, and 1980s. If Snow were to write that book
today (see Price, 1970, for a witty “update”), it would be difficult for
him to avoid making the brain laterality connection. Chapters 12 and 13 are
devoted to "two cultures" matters.
Result‑Driven Thinking
RB‑frontal neural
circuits have many ways of restraining and manipulating LB‑frontal activity.
One of these is called "result‑driven thinking." A thinking process is recognizably
result‑driven when the logical process leading to a position is embarrassingly
contorted and self‑serving, making it obvious that the “result” preceded
the progression of thought that purports to lead to the result. The embarrassment
may be apparent to an objective observer, who is not affected by the position
taken, while for the result‑driven person there is no apparent flaw in a
process he believes to be logical. Deception is always more convincing when
the deceiver is unaware that he is engaged in a deception. Thought blinders
are a useful tool when RB‑frontal wishes to remain undetected by LB‑frontal.
The Mexicans have
a special phrase for calling attention to someone's result‑driven thinking:
quando conviene. Literally translated, it means "when convenient"
‑ which conveys the idea that a person's stated belief changes in a way that
conveniently serves the person's selfish goals. I am convinced that result‑driven
thinking is a "human universal" (see Brown, 1991, for an extensive treatment
of human universals). It probably can be found in all societies and has existed
for a long time.
One of the primary
tasks for result‑driven thinking (RDT) is to force the individual to stay
in loyal service to his group – no matter how illogical. The group may be
his extended family, or his tribe, which in either case is composed of genetically
related individuals. Consider the matter of "tribal mentality," first described
by Spencer (1892) and later by Keith (1946, 1948), which refers to the anthropological
finding that all primitives live by a moral system that requires beneficent
treatment of fellow tribesmen while condoning and even encouraging barbaric
treatment of individuals belonging to neighboring tribes. "Tribal morality"
requires an illogic for which RDT is admirably suited, and for which the
RDT capability may have evolved. Empathy and amity for some, enmity and hostility
for others! This seems at odds with contemporary religious dogma, yet in
time of war both sides appeal to their respective religions for legitimacy
and support in vanquishing the enemy. Each side distorts the character of
the other, making up false accusations and using degrading names for the
purpose of arousing the passions in preparation for combat.
Political partisanship,
ever more prevalent in America since World War
II, also illustrates the power of RDT. (I adopt the common usage of the term
"America" to refer to the
United States, in apparent neglect
of the fact that "America" actually includes
Canada, Mexico, and all of Central
and South America.) The model for
this is tribal conflict. Consider the case of the U.S. Supreme Court's 2000
decision that in effect led to George W. Bush's selection for president.
The 5 conservative‑leaning jurists had a longstanding record of supporting
“states rights” on many matters, while the 4 more liberal jurists favored
federal rights, yet when the issue was the election of either a conservative
or liberal for president both sides switched their positions concerning which
right trumped the other, and by acting in this unusual way they served personal
political philosophies. Chief Justice Rehnquist later alluded to the need
for the Supreme Court to sometimes get involved in political matters to prevent
a national crisis (Los Angeles Times, January 21, 2001, pg. A30). Quando conviene!
My reading of the
newspaper is immensely more informed because I readily recognize RDT. Almost
every opinion, and the rationale for almost every action, requires RDT to
protect it from the detached, logical thinking that a well‑functioning LB‑frontal
is capable of. There is a common misperception that people are LB‑dominant,
which is based almost solely on where language capability resides. But
in the realm of thinking, humans are RB‑dominant. And RDT is the mechanism
for accomplishing this in the presence of a powerful logical left brain.
RDT is a tool of
the genes to have things both ways. By inventing RDT, the genes have become
able to receive the benefits of a powerful new LB while retaining their interests
in enslaving the individual for genetic service. Obedience of the powerful
new LB is achieved by RB circuits that assure that "result‑driven thinking"
is preserved in a way that accomplishes genetic goals.
Thinking is
a Subversive Activity
Occasionally I’ll
buy a book based solely on the cleverness of its title. My favorite example
of this is Teaching
is a Subversive Activity. I never read the
book, probably because I didn’t want to be disappointed. The title by itself
inspired me to develop a text in my imagination, and over the years I’ve
continued to add to that imaginary text. This section will draw from that
text.
Society endorses
a school curriculum that renders students “useful” to society, regardless
of how useful the curriculum is to the student’s individual fulfillment (thanks,
Alfred Allen, for stating it this way to me). If heroism is useful to society,
then history and literature classes will feature heroism. If “slacking”
is not useful to society, then the payoffs for slacking will not be taught.
In between these two extremes are such things as tolerance, celebrating traditions,
skepticism and questioning authority. If a curriculum were to be designed
to serve individual fulfillment it would suggest that heroism is folly,
tolerance is good, skepticism is essential and all authority should be questioned.
The fact that none of these are to be found in schools is unsurprising.
“Thinking” is something
the left brain does. It is also something the right brain “controls.” If
thinking occurs, it is because RB permits it, and may even encourage it because
a problem needs to be solved that appears to be compatible with the genetic
agenda. Thinking is inhibited by RB when it appears to threaten the genetic
agenda.
When a parent enters
a burning house to save a child there is a quick and mostly subconscious
calculation of the danger involved, and the resulting decision favors genetic
interests. When a household is attacked by intruders the ensuing defense is
likely to include heroic acts. When a tribe is attacked by a neighboring tribe,
or when one country is attacked by another, heroism will be found among the
defenders. Each heroic act will be called “selfless,” or what the intellectuals
would call “altruistic,” and we can be confident in surmising that these
acts were initiated by a right brain that was able to inhibit a left brain
from hesitating for thoughtful consideration.
When society pays
a teacher’s salary it is natural for society’s interests to be served by
what’s taught. Any teacher who suggests that individuals have the right to
renounce heroism when a situation calls for it would be considered subversive.
Similarly, any teacher who counsels skepticism and a questioning of authority
would also be considered subversive – especially by the authorities.
America has become a “consumer
society” and the American government has become the protectors of business
interests. Imagine schools that teach how to question advertisements, make
fun of claims by the pharmaceutical industry, question FDA approvals, or
take every politician’s utterance and consider that truth is closer to its
opposite. Students graduating from such a school would fare better in life,
but those with power in society would view the curriculum as subversive and
they would not permit such a school to exist.
In a future chapter
I will describe the co-evolution of genes promoting altruism and intolerance.
A theory will be described that purports to show that when there is chronic
conflict between neighboring tribes there are benefits for the tribe whose
members are both intolerant and altruistic. Heroes will be seen as intolerant
altruists! Only when a tribe overwhelms all competitors
is it safe for genes promoting the opposite traits to prevail. Thus, each
empire will gradually become dominated by selfish individuals who tolerate
others regardless of their differences. This becomes a “weakness” from
the standpoint of societal survival, which leads to the decline and fall
of the empire. Only after this theory has been explained will it be possible
to understand the full meaning of “thinking is a subversive activity.”
─────────────────────────────────
CHAPTER
9
─────────────────────────────────
ARTISANS SET THE STAGE FOR
CIVILIZATION: PART I
The human brain
and culture co‑evolved (Lumsden and Wilson, 1981) during at least the past
70,000 years. In this chapter
I will make the case for the position that a civilization can only develop
when there is a division of labor, and when society supports a large base
of "artisans."
The first artisan
niche was probably the toolmaker, whose task was initially part‑time, but
eventually became full‑time. Significant evolutionary forces were created
when the first full‑time artisan was employed, and these led to the further
specialization of the left brain (LB).
When the Pleistocene
glacial climate began a transition to the Holocene interglacial, 14,650 yr
BP (years before the present), the better climate set the stage for an explosive
expansion of artisan niches. Increasingly complex economies allowed for
higher population densities, which supported large, sedentary populations.
New artisan niches were created, allowing for an increase in the artisan
population. The increasing "presence" of artisans caused cultures to expand,
and become more sophisticated. The artisan played a crucial role in creating
civilizations.
The entire process
of artisan proliferation, cultural elaboration, and the creation of modern
civilizations, occurred because human culture and brain‑function genes co‑evolved.
By this is meant that the civilized environment that was created by artisans,
whose special abilities are at least partly due to the appearance of "artisan
genes," changed the environment in such a way that artisan genes were more
valuable, and were "selected" in greater numbers. This chapter describes
some speculative mileposts along this interesting journey.
Pleistocene Life
I will argue that
for the past 70,000 years one of the most strongly contested allelic competitions
was related to the creation of full‑time niches within human tribes.
The world's climate
was cold for most of the past 1.6 million years, a period referred to as
the Pleistocene Epoch (from 1.6 million to 12,000 years ago). Brief warmings,
or interglacials, occurred at approximately 100,000 year intervals throughout
the Pleistocene. There was an interglacial from about 129,000 to 116,000
yr BP. At about 69,750 yr BP there was a brief several‑century warming, but
it was too dry and not quite warm enough to qualify as a true interglacial.
A brief warming occurred 34,800 yr BP. After an extreme cold period 18,000
yr BP, a gradual warming began. Erratic swings of warm and cold climate gave
way 14,650 yr BP to an almost irreversible warming (the Younger‑Dryas cold
interlude was from 13,000 to 11,600 yr BP), ushering in a true warm interglacial,
called the Holocene Epoch, extending from 11,600 yr BP to the present.
Prior to the “70,000
yr BP warm/dry episode” human tools were uniformly simple. After this warming
event tools became abundant, standardized, and more sophisticated (in Africa and Europe.) What caused the
proliferation of quality tools? One possibility is that full‑time toolmakers
appeared at this time.
In order to understand
the importance of tool making we must imagine what life was like during the
Pleistocene. What, for example, were the main "selecting forces" for humans?
In the book Demonic Males (1996) by Richard Wrangham and
Dale Peterson a compelling and disturbing description is presented of our
ancestors, starting with woodland apes that split from the chimpanzee line
5 to 7 million years ago, then to proto‑humans living from 1.8 million years
ago to 200,000 yr BP, and finally to our homo sapien ancestors of the past
200,000 years. All of these ancestors, like their chimpanzee ancestor, were
preoccupied with territory, border raids, rape and even warfare ‑ accomplished,
of course, by "demonic" males.
A book by Lawrence
Keeley, War Before Civilization
(1996), carries this theme into the late Pleistocene and Holocene. An archeological
record is described brimming with evidence for pre‑historic "wars" that were
more deadly and pervasive than our bloodied modern experience. Women and
children were not spared, except for abduction as new wives or to serve as
slaves. The enemy's property was either destroyed or appropriated. Mass
graves contain victims with embedded arrow heads or spear points. Palisades and fortresses
with moats preceded castles by many thousands of years. The principal cause
of death during the past 20,000 to 30,000 years, when the archeological record
is good enough for these interpretations, appears to have been inter‑tribal
warfare, with surprise raids being the preferred strategy.
Our ancestors must
have lived in a world where survival depended on effectiveness in warfare.
The tools of war during the last 200,000 years of the Pleistocene must have
been clubs, axes, spears and eventually bows and arrows. The term "hunter
and gatherer," in which men hunt and women gather, should be amended to "warrior,
hunter and gatherer." Therefore, our "ancestral environment" (AE, also referred
to by the more pedantic term "environment for evolutionary adaptation,"
or EEA), should be based not on the H&G model, but on a WH&G model!
Colin Tudge (1998)
summarizes emerging suspicions that throughout the past 100,000 years our
ancestors supplemented a "hunting and gathering" mainstay by horticulture
and pastoral "farming." Horticulture consists of weeding and clearing an
area to provide for a greater growth of some naturally occurring plant that
produces something that is normally "gathered." Pastoral farming consists
of herding and later domesticating animals that were previously hunted. Both
forms of "proto‑farming" invite banditry! And banditry will inevitably elicit
defensive efforts by the proto‑farmers. Both bandits and farmers would require
effective weaponry, and this increases the importance of artisans who specialize
in weapon‑making.
Weapon Making Transitions
I speculate that
prior to the 129,000‑116,000 yr BP interglacial, every hunter/warrior made
his own tools ‑ including axes, spears, bows and arrows, which were used
for both hunting and warfare. After the 129,000‑116,000 yr BP interglacial,
a part‑time toolmaker may have made most of these items. But during the 70,000
yr BP dry warming, migrations may have enhanced tribal conflict, rewarding
those tribes with the best weapons. Any tribe that made use of a full‑time
weapons maker would have enjoyed an advantage in inter‑tribal raids. The
critical warming 70,000 years ago would then have completed a two‑phase transition
between three states: 1) a culture in which "every man made his own weapons
and tools," 2) part‑time tool and weapon making for the others, and 3) a
tribal arrangement in which a specialist assumed full‑time responsibility
for tool and weapon making.
Whether the first
full‑time specialist appeared 120,000 yr BP, 70,000 yr BP, or 40,000 yr BP,
the event would have been a milestone in human evolution. With a niche in
each tribe for just one thing, such as tool-making, genes for tool-making
could be evolutionarily selected at a rapid pace. Before that time, any genes
for tool making are likely to have had deleterious effects on other phenotypic
traits, which would have represented a serious penalty. But if a person's
task is confined to mostly tool-making, it doesn't matter if he is unable
to perform as a warrior, for example. Fierceness isn't a requirement for
tool-making, nor is physical strength, fast reflexes, or endurance. A new
era dawned for humanity when this new full‑time niche came into existence.
Because of what followed, I shall refer to the people who filled these "weapon
maker" niches as "artisans."
Weapon Makers, Toolmakers
and Artisans
The first full‑time
weapon‑making "artisan" may have differed little in temperament from his
fellows. It would make sense for him to train an apprentice after achieving
competence, in order to preserve what had been learned through years of trial
and error. This arrangement, in which an artisan takes
on an apprentice, set the stage for the accumulation of advances in weapon
making technique during succeeding generations.
Anyone expert in
making weapons is already more likely than others
to make good tools. The axe that kills an enemy warrior is similar to the
axe that chops a tree. The stone knife that cuts an enemy warrior's scalp
for a take‑home trophy could also be used for skinning an animal. The first
tools for constructive uses could have had their genesis as warrior weapons.
The weapon maker was a natural choice for assuming tribal toolmaking duties
as well.
If the apprentice
is chosen by observation of who seems to have talent and temperament for
toolmaking, and if this practice is preserved for many generations, then it
is inconceivable that the genes would "overlook" this new opportunity. “Toolmaking
genes” would have been rewarded, and they would become more abundant ‑ even
if they meant the individual was poor at hunting and "war making." The artisan
will have been released from the genetic limitations imposed by the need
to preserve hunting and warlike traits. Initially, the niche would be limited
to approximately ~2% of the male tribal membership, but an important process
would have been set in motion.
The toolmaker established
a precedent for special status, and this precedent increased tribal readiness
for establishing niches for other artisan work. Proto‑farming might have
created three other niches for artisans: horticulture, animal herding and
animal domestication. Individuals especially effective in these activities
would contribute to a tribe's success, possibly saving them from extinction
when the vagaries of climate or animal migrations brought hardship to those
who relied too completely upon hunting and gathering. Tribes that "accommodated"
proto‑farmers, in addition to full‑time toolmaking, should have fared better
during the late Pleistocene. (If this sounds like "group selection," it is!
I pursue this further in the next chapter.)
If the horticulture
artisans produced a surplus of crops, and if they could somehow process them
for "storage," then there would be payoffs for the new specialty of preserving
surplus foods and building crop storage houses. Because of the physical
demands of this work, the initial stage of construction was probably performed
by men. Whereas we can easily imagine that horticulture was initially women's
work, since gathering is traditionally for women, the first horticulturists
were probably women. But a woman's labor is finite, and childrearing, regular
food gathering and preparation, making clothing, and other domestic jobs
set a limit on how much of her time could be devoted to new artisan tasks.
Thus, over time, men must have assumed more and more full‑time jobs performing
the horticulture tasks. Men probably were the initial pastoralists, since
working with wild animals would probably be dangerous and require physical
strength.
As artisan niches
expanded, it is inconceivable that genes would not have been affected by
the new opportunities, and they must have responded by "producing" people
who were talented artisans to fill the niches. If 10% of the jobs for men
were artisan‑like, then in a steady state condition it can be anticipated
that approximately 10% of men would be born with a phenotype having the artisan's
talent and temperament. As tribes became technologically more sophisticated,
the spectrum of abilities that people exhibited would have "matched" the
broadening spectrum of niche opportunities.
Dawkins wrote about
the hypothetical case of a population of "hawks" and "doves" living with
specified payoffs (Dawkins, 1976), and he showed that natural selection forces
should eventually lead to the establishment of a specific population mix.
It was stable at this "stable point" because any displacement would tilt the
rewards to those who were less populous. He called this dynamic an "evolutionary
stable strategy," or ESS. The same argument should apply to tribal niches,
and modern societal niches ‑ provided the niches are long‑lived and the forces
of selection are natural. Thus, if a tribe "needs" only 10% of adults to engage
in "infrastructure" matters (building and maintaining huts, clearing paths,
building water storage structures, irrigation, sewage disposal, etc), we
can expect that about 10% of newborns will be talented in these activities
(assuming the payoffs are not drastically uneven). Another solution form,
as Dawkins also points out, is that all newborns will be talented in "infrastructure"
but only 10% of them will adopt that role when they grow up. Perhaps, in reality,
we should expect to encounter something intermediate between these two extremes.
A new complication
arises when within one species two or more "types" of individuals are rewarded.
Since females are the result of breeding with males of the warrior type,
they cannot be expected to produce male offspring of the artisan type without
invoking some additional selection mechanism. One candidate mechanism is to
hypothesize that a new type of woman co‑evolves with the new artisan man.
The artisan wife would have to assortatively mate with the artisan man for
this to work. As Dawkins also explains (1982), daughters of a father who differs
from the norm are likely to prefer men like her father, since she inherited
some of her mother's predispositions. The theory for this has been worked
out mathematically, and it is called "linkage disequilibrium." Therefore,
it may be theoretically possible for boys to be born with a predisposition
to become artisan men as opposed to warrior men, or politician men, etc.,
and still find women willing to marry or mate with them.
A simpler mechanism
is for most of the new artisan genes to be located on the same chromosome,
preferably close together. Then, when the sex cells are created by meiosis,
the crossing‑over process is likely to preserve the association of artisan
genes on the same chromosome segment of the new gamete, and thus pass to
male offspring either an undiluted warrior type or an undiluted artisan type.
Let it be noted,
here, that the artisan performs tasks that require good left brain function.
So when artisan niches expand, this is equivalent to stating that there are
genetic rewards for genes that produce individuals who have especially well
developed left brains. This will become an important point later in this
chapter, and in subsequent chapters.
Problems Created
by the Existence of Artisans
If full‑time toolmaking
led to toolmakers who were exempt from the dangerous
exploits of war, who even began to lose their ancestral adaptation to hunting
and war‑making, then what might have been the attitude of the hunter/warrior
toward the toolmaker? Would they not make fun of the toolmaker for staying
home with the women and children when they went out on dangerous hunting
and raiding expeditions? Would they not be inclined to tease and intimidate
the toolmaker, and steal his provisions? But since the person who makes superior
arrowheads and spears is too important to go on risk‑prone hunts, especially
if he is poor at such things, the tribe would be served well by customs that
honored the toolmaker's special status. By stating that the tribe would be
served well if the toolmaker is somehow allowed to safely pursue his labors
without the threat of harassment by warrior men, I am actually saying that
the warriors would be served well by customs that provide for the toolmaker's
protection. This dilemma might have been solved by a ritualized granting
of special status to the toolmaker, with taboos (eventually converted to "laws")
requiring that his tool works and other provisions be off limits to the destruction
or theft that might have been tolerated for non‑toolmaker victims.
Artisan men may
have been shunned by women, for the simple reason that they can not protect
a wife and offspring from "take‑over" males. Moreover, the tribe might benefit
by artisan men not marrying, for a bachelor artisan would have more free
time to practice his essential trade. (Might it be efficient if the genes
also conferred upon the artisans a predisposition for homosexuality? This
is a completely new theory for the origins of homosexuality.)
An unmarried artisan
would be without the benefits of in‑laws to support his case against unfair
treatment by cheats and bullies, who might covet his possessions, food supply
or hut. Social pressure is an important stabilizing force within a tribe.
When someone cheats another, rumors of the wrongdoing spread, and although
this may not restore equity it might at least serve to discourage a repeat
offense. An unmarried man has half as many people belonging to the "relatives
and in‑laws" category, who stand ready to support
him with social pressure, rumor spreading, or literal assistance. These problems
constitute a challenge to the tribe (or rather the genes within the tribe)
to institute an effective structure of taboos that guarantee protection
of the artisan from non‑artisan men.
There are two ways
to imagine how a protection could be accomplished. Tribes that just happen
to include taboos prohibiting intimidation and theft will prosper more than
other tribes. This is a "group selection" theory for the development of taboos,
and eventually the rule of law. An alternative is to suppose that genes
are created (and are present in the population at large) which predispose
people to respect "fairness." And such predispositions favor the adoption
of specific tribal laws which protect artisans (and are available for dealing
with other specific fairness issues). This is a more "robust" path toward
the creation of laws. It requires the co‑evolution of genes and culture,
dealt with below.
It is possible that
the laws which were meant to protect artisans, whose numbers were surely small,
were made use of to some lesser extent by the others. Although the others
would have had less need to use the laws on their behalf, the opportunity
nevertheless existed for these others to "borrow" the protections of status
meant for the artisans by presenting themselves as having artisan abilities.
(A theory for "status" might be developed from this idea, but not here.)
The responses just
described constitute the beginnings of a new type of culture, one based on
concepts of "fairness." The fairness "culturgen" must have been unfamiliar
when it first became a tribal law ("culturgen" is a term for an element of
culture, introduced by Lumsden and Wilson, 1981). It must have been extremely
frustrating for warriors to resist taking advantage of the artisan. The idea
of "status" was old, but the idea of a special status for an artisan, someone
who could not defend his possessions or wife in the traditional manner,
was new. It would serve as a model for new kinds of status that are indispensable
for a modern civilization.
Further Problems
with the Existence of Artisans
Let us be mindful
of the sobering fact that all of our ancestors before the Holocene lived
in a tribal setting. Tribes flourished or floundered as a group. It would
be amazing if we didn't have many genes adapted to tribal living. The tribe
needed an artisan, and the artisan needed the tribe. The tribe fed him, and
protected him from the cruel, harsh world outside the tribal setting.
It's difficult for
us in this individual‑worshipping, modern culture to imagine how restrictive,
confining and enslaving the tribal setting was, and how important group evolutionary
dynamics were. It is often stated that banishment from the tribe was equivalent
to a death sentence. The kind of liberated, individual‑thinking that we
take for granted today would have been rare for our tribal ancestors. Criticizing
tribal rituals or beliefs would have been unthinkable, unless the individual
was willing to leave the tribe and go live by himself, leaving no progeny,
which is an evolutionary dead end. Maybe some people did this, but none
of them are our ancestors.
In a tribal setting
it makes sense for some of the membership to have assigned roles that contribute
to tribal welfare. Individuals could have performed these roles better without
the burden of family. Hence, bachelorhood (maybe even homosexuality) could
have had a place in the tribal society. To use a recent example, we should
all be thankful that Beethoven wasn't a family man, and that Einstein didn't
allow family responsibilities to burden him. A whole host of other lesser
people could also serve the tribe in this way (as they do in today's society),
and they would be better able to make their contributions by eschewing family
responsibilities.
These people, the
artisans, were expected to make individual contributions to the greater good
of the larger group, and part of their individual sacrifice might have been
to forsake marriage. To the extent that the artisans were expected to remain
single, any young man with artisan abilities would have been perceived by
women as a bad mate prospect. Parents may have steered their daughters away
from men who appeared to be on this path of individual contribution. Tribal
people must have had their terms for geek, or egghead, and they would have
served the purpose of discouraging young women from being attracted to “bad
bet” mates. So, anti‑intellectualism may in fact have its origin long ago,
with the artisan playing the role of today's intellectual, being shunned,
yet valued for the greater good.
In spite of all
the special privileges bestowed upon the lucky artisan, he must have had
many unexpected challenges to his individual welfare. Survival of the tribe
is evolutionarily irrelevant except to the extent that the tribe's survival
was a precondition for the survival of the genes within the individual. Thus,
loyalty to the integrity of the tribe would have been valued by all. But
the artisan is a special case. Artisans in all tribes might have been viewed
as somewhat interchangeable. For example, if one tribe triumphed in battle
over another, they might actually go out of their way to not kill or injure
the enemy's artisan, for they could abduct him, and put him to use back home.
Now, knowing this, every tribe should be suspicious of their artisan's allegiance
to tribal survival, for he would have a less compelling reason for adhering
to such an allegiance. Hence, even in the absence of evidence that his allegiance
should be questioned, the artisan should be a worthy target of suspicion,
and he should be treated as someone prone to tribal disloyalty.
The artisan trade
must have brought with it many risks. Imagine a condition in which one tribe
is being beaten down by a neighboring tribe, and weakening year after year.
Might the artisan want to escape before it's too late, and thus avoid the
risk of having to be captured during battle? He might even be at risk of
being killed by his own tribesmen during their defeat, as a form of "scorched
earth" strategy that is even today sometimes practiced. These conditions are
conducive to all kinds of complicated intrigue, all revolving around the
questionable loyalty of the artisan to the tribe. All artisans must have been
both loved and hated by his fellow tribesmen. During battle, indeed, they
would want to protect him because he makes their arrowheads. However, he would be the first to defect in the face of
a deteriorating tribal situation, for he would be accepted by any other tribe.
Oh, how his fellow tribesmen must have loathed his envious position.
Alas, the modern
intellectual is heaped with the same scorn and ambivalence. During WWII the
scientists and engineers who powered the war machinery were the artisans
of their day. Britain’s Alan Turing played a crucial role in breaking the
code of the German’s Enigma Machine, and helping the war effort immensely
(interestingly, Turing was “gay”). After the war Werner von Braun was snatched
by the Allies as if he were a prized booty of victory. How ruinous it would
have been to the Axis if the Allies had captured von Braun at the beginning
of the war. The Germans were short‑sighted to allow Albert Einstein to leave,
and a few others, who contributed to the conception and building of the atomic
bomb. We may never know if Heisenberg intentionally did not work diligently
to build the atomic bomb on Germany's behalf. Such
is the power of the modern artisan, for if Heisenberg had pursued the atomic
bomb successfully, Germany could have easily
won the war.
Returning to the
tribal setting, think of how the artisan must have viewed his fellow tribesman.
The warriors he must have viewed with disdain, for couldn't they see that
the warrior was mainly “fodder” for useless battles (that settled nothing
as far as the artisan was concerned). The women who willingly became burdened
with child‑bearing, couldn't they see that they were being used for the tribal
goal of producing warrior fodder?
And what about the
tribal chief, who protected the artisan from exploitation by the more savvy
and intimidating warrior?
The chief was the artisan's benefactor, so the artisan would at least
have to pretend to view him with a more loyal heart. But the chief must have
thought of his artisans as a "useful asset" ‑ like a herd of cattle, needing
protection in the same way as a cow or goat. The king must have secretly
snickered over this person unworthy of battle, inexcusably effeminate, but
also essential for tribal survival, and contributing to the chief's job security.
So the chief must have had to control his ambivalent feelings toward the
artisan better than the other tribesmen.
The artisan does
not completely belong to his tribe. Rather, you could say that he belongs
to his trade. For whichever tribe fails him, his trade will remain as his
means for livelihood. The artisan secret motto might have been "if you're
good at your trade, the tribesmen will come a courting."
The Holocene Artisan
Explosion
The 5000 year climate
transition from 18,000 to 13,000 years ago was a watershed period for human
evolution. For the first time very large tribes assembled
and adopted sedentary lifestyles. As glaciers receded they exposed
new fertile lands, bathed in warmth and rain, and farming became more feasible.
The domestication of both plants and animals was practiced more widely. Artisan
opportunities exploded, expanding from weapons and toolmaking to such novel
things as animal breeding, irrigation, grain storage, record keeping, trading,
and tax collecting.
The Holocene should
be viewed as an epoch of food surpluses created by sedentary economies, driven
by the dramatic expansion of new artisan niches, which in turn created ever‑more
artisan niches. This positive feedback dynamic fueled an explosion of cultural
change, as well as an exploding population. Large population centers influenced
farming practices across ever‑larger surroundings.
Many aspects of
the way humans lived underwent dramatic change during the early Holocene.
One that deserves comment here is that social life for the first time faced
the challenge of having to deal with strangers who were not enemies from
a rival neighbor tribe. Indeed, some of the strangers encountered in everyday
life might have come from tribes that used to be rivals, but who could no
longer be treated as enemies since they were a useful part of the expanding
new economy.
How confusing it
must have been for the first super‑tribesmen: they were surrounded by unfamiliar
faces, yet these unknown faces were not the enemy. What profound implications
this must have had! Aggressive behavioral responses that were meant for strangers
must have been triggered at subconscious levels, almost continuously, for
early Holocene man while he conducted commerce on busy city streets among
strangers engaged in a similar commerce. The new conditions of public life
called for a change in one's attitude toward "society," as well as one's
relation to it.
The glue that held
together tribes numbering in the hundreds, as with smaller primitive societies,
was based on "inclusive fitness" relationships and repeating "reciprocity"
dealings with familiar tribesmen whose history of faithful past dealings
was known. The new social setting required a greater adherence to explicit
"rules" ‑ which resemble taboos. Concepts of "fairness" were changed, as they
included "outsiders" for the first time. Barter of goods for goods, and goods
for services, and services for goods, became an everyday way of meeting needs.
Artisans, who worked with their brains instead of their brawn, were a newly
respected class. Whereas perhaps 2 to 4% of the pre‑Holocene tribe was an
artisan, perhaps 10% or more of an economically connected population were
artisan‑like. Artisan types proliferated; instead of just toolmakers, the
new artisans constructed irrigation works, farmed, processed food, stored
grain, tended markets, made clothes, kept records and governed.
What had happened
to the old structure, with just hunter/warriors and maybe a part‑time toolmaker?
Few people hunted, and the warrior class had shrunk to a minority, with diminished
power. Things had been turned upside down during the hectic few millennia
spanning 13,000 to 6000 years ago.
This was the transition
to a new condition called “civilization.”
─────────────────────────────────
CHAPTER 10
─────────────────────────────────
ARTISANS SET STAGE
FOR CIVILIZATION: PART II
The Co‑evolution
of Genes and Culture
What is culture?
Is it created by the genes, or superimposed upon human behavior from the
"outside" as a new environment within which the genes must adapt and with
which the genes have no "experience"? Or is culture something in between?
Let's begin with
an 1896 assessment by H. G. Wells of what culture is and why it is so important
to the Human future.
"...it appears to
me impossible to believe that man has undergone anything but an infinitesimal
alteration in his intrinsic nature since the age of the unpolished stone. ... A decent citizen is always controlling
and disciplining the impulses... ...it is indisputable
that civilized man is in some manner different ... But
that difference ... is in no degree inherited. ...
With true articulate speech came the possibilities of more complex co‑operations... Came writing, and therewith a tremendous acceleration
in the expansion of that body of knowledge and ideals which is the reality
of the civilized state. ...in civilized man we have (1) an inherited factor,
the natural man, who is the product of natural selection, the culminating
ape, and a type of animal more obstinately unchangeable than any other living
creature; and (2) an acquired factor, the artificial man, the highly plastic
creature of tradition, suggestion, and reasoned thought. ...in a rude and
undisciplined way indeed, ...humanity is even
now consciously steering itself against the currents and winds of the universe
in which it finds itself. In the future, it is at least conceivable, that
men with a trained reason and a sounder science, both of matter and psychology,
may conduct this operation far more intelligently, unanimously, and effectively,
and work towards, and at last attain and preserve, a social organization
so cunningly balanced against exterior necessities on the one hand, and the
artificial factor in the individual on the other, that the life of every
human being ... may be generally happy. To me, at least, this is no dream,
but a possibility to be lost or won by men, as they may have or may not have
the greatness of heart to consciously shape their moral conceptions and their
lives to such an end." H.
G. Wells, "Human Evolution, An Artificial Process,"
Fortnightly Review, Oct, 1896.
Wells viewed human
nature as unchanging during our acquisition of a changeable culture. He leaves
unaddressed whether or not he thought cultural changes were influenced by
the genes (which hadn’t been generally recognized in 1896), but he doesn't
believe that culture changed our inherited nature (our genotype).
Charles J. Lumsden
and Edward O. Wilson, in their book Genes, Mind and Culture
(1981), take the position that the evolution of culturgens causes evolutionary
change in the genome, which in turn allows new directions for culture,
etc. Each affects the other, and together they co‑evolve, reaching the
elaborate cultural level of today's modern world. Their book is excessively
mathematical, and can discourage all but the most accomplished mathematician;
however, it is not necessary that one follow the rigorous mathematical treatments
in the book to comprehend the concepts presented. I agree with their argument,
and will present a synopsis of it here.
It is easy to understand
that cultural evolution is channeled by what is "possible." As a trivial
example, consider a hypothetical cultural element, or "culturgen," forbidding
the drinking of liquids. Any person fool enough to adhere to this regimen
would die (unless sufficient liquids were present in the solid diet). Not
only would practitioners not live long enough to spread the culturgen, but
the need for liquids is so strongly rooted in our genes that the weird culturgen
would lack appeal and fail to win converts.
A pork taboo, on
the other hand, would face less resistance, and indeed has appeared at a
location and time when eating pork was probably too risky to be worth its
nutritional benefit. Thus, the genes "allow" some culturgens but not others.
The incest taboo
is a well‑studied culturgen, and it is found in all societies. Genes have
evolved that identify incest situations and produce an aversion to their
completion (in order to prevent the homozygotic expression of recessive genetic
defects in offspring). The incest taboo is present in all human societies
as well as many species. When it appears in animals that we normally do not
believe are capable of culture, the behavior can be said to be hard‑wired.
It is not strongly hard‑wired in humans because incest behaviors do sometimes
occur, and details of the taboo are different in different cultures.
Incest taboo is
a "permitted" culturgen; indeed, it is a predisposition that the genes have
been coding for during all of human and pre‑human ancestry. Therefore, any
genes that influence incest behavior will be under selective pressure, and
can be quickly selected into existence if they exhibit adaptive nuances ‑
such as favoring first‑cousin matings.
The following hypothetical
culturgens illustrate the range of likely to unlikely: 1) the celebration
of successful warriors versus scorning them, 2) offering help to fellow tribesmen
versus offering help to neighboring tribesmen, 3) sharing food with relatives
when there's extra food versus denying food to relatives, and 4) adopting
tribal culturgens versus mocking them. The genes aren't "dumb," and not
all culturgens have an equal chance for acceptance.
The "other side
of the coin" is to ask if the evolution of genes can be affected by an entrenched
culture? The key word here is "entrenched." There
is a tendency for all members of a tribe to adopt the same culturgens, a
noticeable human trait called "conformism" (Boyd and Richerson, 1996; Henrich
and Boyd, 1998). In sharing a culture, people adopt most of its culturgen
elements. There must have evolved a gene for a brain function that causes
individuals to be unquestioning joiners, and all people have the gene.
Consider a person
who is more willing than others to try out new culturgens. Not only will he be burdened with many new culturgens, most
of which will be maladaptive, but his beliefs and behaviors will make him
resemble someone from another tribe. Since every tribe is in conflict with
neighboring tribes, a person who appears to belong to another tribe will
be severely handicapped in gaining acceptance by his own tribe. Unless this
open-minded individual happens to adopt a highly adaptive culturgen, his
aberrant beliefs and behaviors will not be tolerated by his fellow tribesmen
and he will be banished by them. Therefore, any gene that inclines a person
to be open-minded is likely to quickly disappear from the human genome.
Yet, new culturgens
do occasionally appear. So there must be a mental
calculus of perceived benefit versus cost that allows some individuals to
adopt a new culturgen without being banished. Perhaps tribesmen who have attained
a position of unquestioned tribal loyalty are able to try out mildly new
culturgens with impunity. If a successful huntsman uses a new arrowhead shape,
then other tribesmen may be curious about it and may eventually adopt it.
If the chief attributes a tribal victory to a new spirit, the others may
consider accepting this new spirit. Everything new has a barrier for acceptance.
If enough individuals are willing to overcome their natural resistance to
a new idea then a generally-accepted culturgen shift within the tribe might
eventually occur. If this happens, then those who remain uncomfortable with
the new culturgen would be at a disadvantage, and the gene that codes for
their culturgen preference would face a slow extinction.
This illustrates
how culture may influence the evolution of genes.
To cite a specific,
hypothetical example illustrating the co‑evolution of genes and culture,
imagine the first groups of Africans to migrate northward after the start
of an interglacial warming. Upon reaching Europe, the migrants would
have encountered retreating glaciers, rivers of melt water, abundant plant
growth, grazing animals, and slow‑moving Neanderthals (distantly related
to humans). These migrations might have occurred 120,000 years ago, 70,000
years ago, and 13,000 years ago (but, in this last case, the migration
would not have encountered Neanderthals, who were displaced by humans about
30,000 years ago). The new setting presents many opportunities, but it
also demands many behavioral adaptations. Seasons are more extreme, and
procuring food in winter is different from summer. Uneaten food doesn't
spoil as quickly as in the jungle, so food storage is not only possible,
it is essential to avoid starvation in the winter.
In this new land
with seasons it makes sense to establish a home base in the fall where food
provisions can be kept for use throughout winter. New customs are needed,
as are new instincts. Whereas jungle life has no rewards for those who store
food, glacier's edge life demands it! The impulse to eat whatever food is
present is now a liability. Impulse control on this, and other matters, is
important. Conscious thought is brought to bear on such tasks as providing
food stores for the winter, protecting these stores from theft by animals
(and other human groups), planning ahead by making winter clothes from animal
skins while the skins are available, finding a cave before others, constructing
a shelter, and many other season‑related tasks.
As described earlier,
every population of individuals will exhibit a spectrum of pre‑adaptations
and pre‑maladaptations to a totally new challenge. Those who are naturally
inclined to possess impulse control, for example, will be inclined to adopt
culturgens requiring impulse control. In the absence of a large tribe with
an entrenched culture, individuals are freer to discover their innate "usage
probabilities" for new culturgens. The transition from a previous culturgen
to a competing new one, is set by the genes, but is also dependent on the
situation (physical environment, social setting, etc). People in the same
situation will have different "transition probabilities." Those who are quicker
to make the transition to the new culturgen are relatively "pre‑adapted"
to the new setting.
Whereas such pre‑adapted
people did not have a competitive advantage in the jungle, they are the new
winners in a mid‑latitude setting. Their pre‑adapting genes create more successful
individuals, and their genes will spread through the gene pool of those
groups that migrate north.
As a new collection
of culturgens accumulate, creating a new culture, some genes become mal‑adapted
(to the new culture). For example, genes for impulsivity handicap individuals
with that gene. If those people fail to set aside winter food stores, and
are forced to steal from neighbors during the winter, they are at greater
risk of injury or death by those who are protecting their food stores. Agreements
may be formulated among like‑minded provisioners, requiring the group to
take action against those who don't respect other people's "property." An
individual who has trouble grasping the new concept "property" and "property
rights," and the consequences of "stealing," will be dealt with harshly by
the majority, once these new culturgens are adopted.
These examples illustrate
how a new environment can change culture, and how a changed culture can influence
the fate of genes, causing gene allele frequencies to change. As one change
becomes established, new selection pressures exist on the other. And selection
pressures work in both directions: new genes alter "transition probabilities"
for the adoption of new culturgens, and newly adopted culturgens alter the
selection pressure on genes. Thus, genes and culture
co‑evolve.
I claim that when
tribes began to subsidize the full‑time employment of artisans, possibly
during a warming 70,000 years ago, the stage was set for an explosion of new
artisan‑like niches, and that when the Holocene interglacial began, some 12,000
years ago, the explosion of changing cultures began. The new niches include
such things as agricultural farmer, domesticated animal farmer, tribute record
keeper, clothier, entertainer, priest, government administrator, entertainer,
merchant, full‑time soldier and others.
Before these changes
began everyone within a tribe assumed the same roles, which dealt mostly
with providing food and fighting neighboring tribesmen. The Holocene saw fewer
and fewer people engaged in the traditional, all‑roles lifestyle; an ever‑increasing
fraction of people in urban centers became engaged in specialized roles,
having nothing to do with food production or fighting wars. Country living
yielded to living within or close to cities. Culture became more complex,
powerful, and played a growing role in the selection of gene alleles that
were pre‑adapted for new niches. This is the story of the expansion of culture
and the birth of civilizations.
Defining Civilization
Let us ponder the
term "civilization." Like most people, I know it when I see it, but it may
nevertheless be instructive to struggle with defining it.
Perhaps the root
word "civil" is the key to its definition. Where civil social interactions
are common, there resides a civilization. However, uncivilized primitive
people are usually "civil" to each other ‑ but their "civility to strangers"
is another matter? It is well known that the "tribal mentality" (Spencer,
1892) requires that two separate codes of morality be used; one is meant for
intra‑tribal interactions (amity) and the other for extra‑tribal interactions
(enmity). However, when neighboring tribes trade goods, they are civil with
each other. Even if that's due to a fear of retribution, fueled by not knowing
the ferociousness of the stranger tribesmen, "civility to strangers" still
lacks the essential trait we're looking for.
I'm going to suggest
a definition based on an observation that has probably never been suggested
before. I assert that a civilization is the product of left‑brained values
and productive activities. Consider the dictionary definition: civ·i·li·za·tion, n. 1. An advanced state of intellectual, cultural,
and material development in human society, marked by progress in the arts
and sciences, the extensive use of writing, and the appearance of complex
political and social institutions. Note how left
brained these qualities are: material and intellectual developments, writing,
science, complex social political and institutions. These
are things that left brains value and only they can do!
The dictionary's
phrase "the appearance of complex political institutions" conceals a deeper
truth about civilizations. One of the functions of complex political institutions
is to safeguard the rights of individuals from violation by the collective.
I believe that ever since the left brain began its specializations for what
we now recognize as LB‑style thinking there has been a conflict between obligations
imposed upon the individual by the group versus LB‑style individual aspirations.
The group wants conformity, and it endeavors to suppress individual expression.
Those individuals who identify with group conformity are agents of the collective
will, which is to say, they are dupes of the genes! For it is the individual
with an independent will, fortified by a strong LB, who is unwilling to
remain subservient to its RB, who can show the way to liberation from the
genes that wish to keep us dedicated to serving their "needs." LB‑style individuals
protect themselves from exploitation by the genes by constructing political
institutions, such as a legislature, a police system, and a justice system.
When these institutions work, they control the collective's meddlesome intimidation
of the individual who is minding his own business as he creates his own individual
path through life.
Too often the political
institution is hijacked by the enemy of the individual. Communism is a case
of the collective usurping power from those who created the institutions
to protect the individual from the insidious meddling of the collective. I
believe that communists are well‑meaning, but misguided by a naive understanding
of human nature. They mistake society for the family. Within a family it is
natural for each to take from those who have and give to those in need. This
makes genetic sense, since the members of a family have a strong genetic relatedness
to each other. It even makes sense for a tribe to behave in this communistic
way, though to a lesser extent than for the family. But human nature will
not endure the attempt to bestow family and tribal obligations to the larger
social group of a society. Society can never be made to look like a family,
or small tribe. Human nature has been molded for competition with neighboring
groups, and it mobilizes our energies to defeat them. Communism must suppress
individualism; it symbolizes the classic conflict between the needs of the
collective and the aspirations of the individual. Communism is the enemy
of everything valued by LB. LB wishes to liberate the individual from the
collective, and communism thwarts these liberating ambitions.
The dictionary definition
also refers to progress in the sciences. Science is a discipline that requires
strong LB involvement. For a scientist, RB must play a supportive role, though
it does "point the way" and "give opinions" when its intuitive feel for matters
is useful. During my practice of science (in the physical sciences) my RB
contributions have been important, but supportive. I will share credit with
RB for my four patents, and other creative labors. Intuition is an essential
guide through the labyrinth of possibilities faced by a researcher in any
branch of science. Hunches that pay off advance every investigation. But
the entire enterprise is overseen and guided by a disciplined LB. After the
inspiring moment (which has happened to me many times), while the emotional
excitement swells, LB goes into action and begins to "work out" the idea.
Logical consequences of the idea are pursued, and tests of it are devised.
A moment's inspiration can lead to many years of an unfolding, LB‑guided
investigation. Without a specialized LB, science could not progress.
As an aside, I believe
the inclusion of "art" in the dictionary definition is a mistake. If we somehow could remove all the "arts" from Western
civilization, would we still think it was a civilization? Imagine that we
had the same level of literature, science, technology, musical heritage,
material standard of living, sophisticated governing institutions, medical
knowledge, and insight into how things work, I
claim that we would still call it a civilization. Primitive societies have
their "art" ‑ and sometimes it's quite good art, easily rivaling "modern
art" in appeal. Cro-Magnon artistic renderings are impressive, yet they did
not possess a civilization. I maintain that "progress in the arts" is not
an essential aspect of a civilization.
The dictionary definition
for civilization refers to "the extensive use of writing." Writing is an
LB activity, with key roles for both Broca's and Wernicke's areas, and others
within LB. True, RB plays a role, but it is a supporting role, similar to
the role I described for the pursuit of science. If RB tried to write by
itself, it would lamely produce pat sayings, open interpretation poetry,
and profanity; it would be unable, for want of a Broca's area, to produce
syntactical prose. This much we know from brain research, that reveals what
RB is capable of verbalizing when LB has been disabled.
There are more candidates
wishing to be called a civilization than are deserving of it, the way I prefer
to define "civilization." For example, the Mayan is often referred to as
a civilization. The more we learn about it, the more despicable
it appears to have been. For every trace of accomplishment, there are several
barbaric, bloody practices. Yes, their artisans devised a complex calendar,
and massive stone pyramids and temples, but they were used for the most inhumane
ceremonies in the human record. The Mayans are an embarrassment to humanity!
The Mayan individual was a victim of his culture (I will grant them "culture"
status, but not civilization status). I pity those poor LB‑style Mayan artisans
who must have existed, for their labors were used to advance a collective
appetite for brutality that only crazed contemporaries could admire. It appears
that the insane Mayan culture was the captive of a right brain that was answering
every call of the reptilian brain. Anyone with a strong LB would have had
a limited opportunity to influence societal values, and would have been relegated
to improving the calendar, overseeing the construction of killing temples,
and fiddling with hieroglyphs for recording the glorious deeds of their murderous
employer.
Most so‑called civilizations
are a mixture of the Mayan type (regrettably dysfunctional) and the ancient
Greek example (admirable). The early Greek civilization produced truly ground‑breaking
insights into the nature of reality and Man's place in it. I will not present
a systematic listing of civilization candidates, and their salient features.
Rather, I will use three civilizations from among the many to illustrate
dynamics that to some extent must have been present in them all. Let us first
consider the rise of one of the first civilizations, the Minoan, and try
to learn what drove its ascent.
Civilization Growth
Phases
The Minoan civilization
grew through three stages: an Early Period, 3000 to 2100 BC, a Middle Period,
2100 to 1600 BC, and a Late Period, from 1600 to 1326 BC, which came to an
abrupt end when the volcano on the island of Thera erupted and, by
destroying much of their infrastructure, rendered them helpless against
invasion by the Mycenaeans. A tsunami probably destroyed the coastal settlements
along Crete's north shore,
where the Minoan civilization was also present. Prior to their demise, the
Minoans were merchants and traders. They plied the Mediterranean, moving products
from port to port for profit. Their merchant ships were well designed, and
very functional. They had multi‑story residences, with water delivered to
some of them, aqueduct style. Their standard of living
must have been one of the best for their time. The Minoans were a peaceful
people, as they apparently had no army. Their art was elaborate and accomplished,
and when the subject matter included humans it was usually related to athletic
performances, dancing, and never warriors, battle scenes or angels.
Although I do not
consider "art" to be an essential component of civilization, it can be used
to understand something about the nature of their society. Art can also be
used to provide clues to the evolution of their rise to power. Consider the
samples of pottery from each of the three periods, and note the style they
used to form and decorate them.

Figure 10.1 Samples of Minoan
pitchers and vases from the Early, Middle and Late periods. From Time‑Life Books (1975).
Ask any neuropsychologist
to view the above sequence of vases and he would immediately recognize that
there's a progression from a functional form preferred by the left brain
to a decorative form preferred by the right brain. LB prefers straight lines
and functional shapes; RB prefers curved lines, ornamentation that is elaborate,
bizarre and sometimes incongruous (i.e., like the baroque
style) and extra flourishes that may detract from functionality.
What could this
progression of patterns mean? Assuming that artisans made what their patrons
wanted, it means that people in power during the Early Minoan Period were
LB‑styled, whereas by the Late Period the power had shifted to RB‑style people.
And, assuming that this interpretation is correct, how could this factoid
illuminate our understanding of how civilizations ascend? It says, I believe, that the earliest stage of a civilization's
rise is driven by LB‑style people. And it also says that during the unfolding
of a civilization the reins of power are captured by RB‑style people. This
last speculation will be taken up in the chapter that deals with the decline
of civilizations.
In Chapter 15 I
present evidence that the per capita output of technological innovations
rises over time to a peak, then subsides ‑ while the economic activity of
the civilization continues toward a peak that occurs a few centuries later.
In the case of the Greco‑Roman civilization, the population peak (a proxy
parameter for economic activity) followed the innovation peak by 5 centuries.
In the case of the present Western (European‑American) civilization, the population
peak will follow the innovation peak (which occurred in 1900 AD) by at least
a century, and probably two centuries.
The innovation peak
corresponds to a period when society gives the greatest freedom to LB‑style
people, by celebrating their efforts, paying for their services, giving them
a status that exempts them from warrior service, and publicly recognizing
that LB activities are good for the general welfare. As I argue in the next
chapter, RB‑style people are "people‑oriented" as opposed to "artifact oriented,"
and they are good at manipulating other people for their personal gain. This
talent of one segment of the population leads to a gradual displacement of
the LB‑style people from power, thus explaining the shift in preferred art
form during the course of a civilization's unfolding.
The world's innovation
per capita has two major peaks, one at 300 BC and the other at 1900 AD. We
know more about the recent peak, so let's consider it from the standpoint
of LB versus RB. It is generally recognized that 15th and 16th Century Renaissance
led to the 17th Century Enlightenment, which led to the explosion of 18th
and 19th Century industrialization. The Enlightenment was a unique chapter
in human history, generated by a changed "climate of opinion." The intellectual
atmosphere was dominated by thinkers who, like Voltaire, penetrated the
cobwebs of previous centuries and saw things the way they were. Voltaire
was a nuisance to the church, politicians, and traditional intellectuals
because he would not be tamed. He saw through the posturing and pretense
of phony pontifications and despised the veneer of social acceptability;
instead, he was cynical, skeptical, uncompromising, and had an acerbic wit.
He exemplifies the LB‑style artisan. Other Philosophes,
like Holbach and Diderot, worshipped the Goddess of Reason, and ushered in
the view that it is within human power to create a world, based upon Reason,
to replace the old unrealistic dream of a Heavenly City, where perfection
and felicity were supposed to dwell for eternity (see The Heavenly
City of the Eighteenth‑Century Philosophers by Becker, 1932). The Philosophes promised a new Heavenly City, built on Earth
by what can now be seen as LB insights, and guided by LB logic.
Civilizations Falter
The 19th Century
began to make good on some of these promises. Inventions just kept coming,
insights into physics accelerated, and Darwin presented the world
with one of mankind's greatest insights into where we came from and who we
are ‑ all based on LB observation and reason. The pace of discovery and industrialization
continued into the 20th Century, starting with Einstein's succession of
profound insights into the nature of the physical world. Science, technology
and engineering were held in high public regard ‑ until the Great Depression.
For a decade, during the global depression in both Europe and the U.S., a malaise stifled
the spirit, and it questioned LB's warrant for carrying the Torch of Progress.
Criticism of society was suddenly unwelcome (poor H. L. Mencken lost audience).
For 15 years serious thinkers were contemplating the end of civilization,
as Diderot and the other Philosophes had worried might happen
during the 18th Century. World War II resurrected the reliance upon technology
for weapons, and the engineer was again cast as savior. The atomic bomb kindled
the bittersweet importance of science. Sputnik forced a new dedication to
giving power to the scientists and engineers, not because of a desire to
build that dreamed of Heavenly City on Earth, but out
of concern for national survival.
The Apollo program
that landed 12 men on the moon was LB's last hurrah! From the social upheaval
of "The Sixties" came a change in the "climate of opinion" ‑ which lasts
until today. LB accomplishments didn't stop, and in fact continue to be made
use of, but they were not publicly applauded (except for an occasional rover
on Mars or Hubble Space Telescope picture). Political correctness was created
to discredit and stifle LB values. The greatest insight that Mankind has
achieved occurred midway through the last half of the 20th Century, yet almost
no one knows about it today. Mankind's greatest discovery is Sociobiology!
It is the crowning achievement of LB thinking. It is comparable to the 19th
Century's discovery that the physical world is reducible to the invariant
laws of nature. Sociobiology forces living systems into this physical world,
and accomplishes the supreme feat of Reductionism ‑ everything, including
life, is governed by invariant physical laws, and all happenings reduce to
an unfolding of physics, where a = F/m and
quantum physics determine everything!
My measure for a
civilization is that people have an honest understanding of who they are.
If only a few percent have this glimmer of understanding, it constitutes
a civilization. The Greeks qualify, thanks to such luminaries as Thales of
Miletus, Anaximaner of Miletus, Democritus of Abdera, and some of the ancient
Romans qualify, thanks to thinkers like Lucretius. The 20th Century, Western
Edition, qualifies because of such sociobiologists as W. D. Hamilton, G. C.
Williams, Robert Trivers, Edward O. Wilson and Richard Dawkins.
The Chinese have
seen many civilizations rise and fall in their land, and during the past
millennium they have often been more advanced than their contemporary European
civilization on measures commonly used to describe civilization. Their technologies
raised living standards, but as far as I can determine they repeatedly failed
the boldness test concerning the quest of insight into the nature of reality.
Their "philosophy" suffers from an excess of intuitive, RB style nonsense!
In my opinion they never achieved the level of insight of Thales, Democritus
or Lucretius, and their 19th and 20th Century stifling
"collective versus individual" culture has made them bystanders while Western
thinkers explored beyond the Greek giants, led by Schopenhauer, Bertrand
Russell, and the sociobiologists.
On many occasions
the Chinese abandoned their relatively advanced technology, and reverted
to living in an RB world. This "failure of nerve," or unwillingness to pursue
Truth into areas where it "hurts," constitutes what seems to me to be an
endemic Oriental flaw. Their practice of physical science suffers from the
same intellectual timidness. Even though the Chinese score higher on IQ tests
than all other races (except the Jews), there's something about their frontal
lobes, or something about their RB‑style of thinking, that causes me to question
their ability to boldly advance human understanding of big picture matters
on "the nature of existence" or "who we are." Until they allow the individual
more freedom from the will of the "collective" the contributions of the Chinese
civilization will be confined to mostly engineering. This, I'm sorry, is
my humble opinion.
Thanks to a specialized
left brain, two great civilizations, by my reckoning, have arisen during
recorded history. I rank them "great" because they celebrate the individual,
and they bring us closer to a stage of human evolution when we shall subdue
the collective mentality. Outlaw genes created this desire to conform to what's
good for the collective good, and we are now discovering that we have been
dim‑witted slaves too long! LB is leading the way to emancipation, and it
is accomplishing its feat by creating civilizations. The job of liberation
has not been accomplished; but the stage has been set for it's serious pursuit.
Thus we stand at
the cusp of two millennia, looking back at many failed civilizations, at
least two great ones, and wondering where ours is headed. Most people are
unaware of human servitude to the genes and the collective they've created;
while others, like me, wish for liberation and wonder if this next will be
the century when humanity's emancipation will finally be achieved. If my claim
is true that each civilization is the result of LB efforts to improve the
life of the individual, with the unforeseen consequence of bringing the individual
closer to liberation from the grip of outlaw genes, then we have a tool for
discerning the health of today's civilization, and predicting its future.
─────────────────────────────────
CHAPTER 11
─────────────────────────────────
LESSONS FROM SAILING
SHIPS:
AN INTRODUCTION TO GROUP SELECTION THEORY
“For now I see peace
to corrupt no less than war to waste.” John Milton, Parasise Lost, 1667
Imagine being a
crew member on a merchant ship setting sail for a crossing of the Atlantic Ocean during the 18th Century. There will
be storms and the constant threat of pirates during the 7-week journey. The
sailing is sponsored by merchants who want the cargo to arrive safely, the
ship’s owner who wants to preserve his investment by the arrival of his ship
intact, and the captain and crew who wish to arrive safely where they will
be paid and continue their lives. All factors favor cooperation by everyone
on the ship in the mission of operating the ship properly on the high seas
and delivering its precious cargo safely to the opposite shore.
Each person on the
ship has one or more assigned jobs. Presumably the assignments are made on
the basis of ability for the needed tasks. It won’t matter that one crew mate
is an excellent runner, or hunter, or mountain climber, or jungle explorer,
for on the ship these abilities don’t matter as he will be measured by his
performance of assigned tasks. Each crew member’s fate will be affected by
the quality of his crewmates and the manner in which they all work together
to navigate the ship safely to port. When each mate discharges his task with
competence and cooperation the entire endeavor is helped, and the prospects
for a prosperous outcome for all mates is improved
This situation is
a simple way to introduce the concept of “group selection theory.” During the voyage all people aboard the ship will either
live as a group, or die as a group.
This is a more extreme example of a tribe either entirely living or dying
during conflict with a
neighboring tribe, but the concept is easier to grasp using the sailing hip example because the ocean is deep and unforgiving with
a history of taking entire crews to the ocean bottom.
With the ship analogy
in mind let’s consider the tribal situation; after all, the tribal setting
our ancestors had to survive for millions of years. If a tribe is in chronic
conflict with a neighbor tribe the losing tribe might be decimated. This
prospect has a message for individual members who pride themselves as being
proficient in some irrelevant realm. An individual with a talent for basket
design, for example, will have a useless talent when there are more compelling
needs for warrior talent.
So what makes a
good warrior? There are the obvious factors of strength, agility and
other skills. Two other
factors deserve special attention: altruism and intolerance.
Altruism is defined
as a willingness to forego individual payoffs in order to achieve a payoff
for another individual or group of individuals. Two explanations are commonly
offered to account for the existence of altruism. First, if the cost to the
altruist is small, and the benefit to the other person is great, and if
the interactants have recurring relationships, then it is easy to imagine
that a series of such acts can yield benefits to all participants if there
are several such interactions with opposite sign. (The “sign” of the interaction
refers to which person is the recipient of the altruistic act.) Notice that
this dynamic does not require that the two people have a close genetic relationship.
The second explanation
for altruistic acts requires that the two individuals be closely related.
J. B. S. Haldane famously quipped that he would willingly give his life for
two first cousins, or four second cousins, etc., in answer to a question
about altruism. The calculus of genetic payoffs of this type is now called
“inclusive fitness” and it states that our brains are designed to recognize
when a sacrifice is likely to confer a greater benefit than loss to our genes,
present in our near relatives as well as in oneself.
Finally, there’s
a “group selection” theory that can account for altruistic acts. If a tribe
is at risk of being decimated by a rival, and if the home tribe is desperate,
then there’s logic in some individuals making high risk attempts to turn
the tide of battle. It’s not necessary for the hero to be closely related
to his fellow tribesmen since all of them will either survive or be killed
depending on the outcome of the battle. This is analogous to ship mates dealing
with an emergency at sea which requires heroic action to save the ship and
all its crew. The genetic relationship of the sailors is irrelevant to the
need for action.
A heroic warrior
can be seen as an altruist. He risks his life in order to save the tribe
because saving the tribe also saves the hero. Genes that predispose to this
form of altruism should be selected for by evolution whenever tribes live
in chronic conflict with their neighbors. The prediction is borne out, at
least in game theory simulations (Choi and Bowles, 2007). Since the altruistic
acts benefit only those in the home tribe it has been referred to as “parochial
altruism” (“parochial” refers to a concern that is narrowly restricted, or
a way of thinking that is “provincial”). The notion that genes predisposing
for “parochial altruism” will evolve when tribes are in conflict is based
on “group selection theory.”
There’s an interesting
aspect to the way in which this kind of parochial altruism is elicited, which
has also pointed out by Choi and Bowles (2007) as well as Wilson and Wilson (2007). It pertains
to intolerance, an unwillingness to overlook individual or group differences.
For example, if fellow tribesmen dress one way and someone is seen dressing
another way (not incorporated into tribal rituals), the non-conformist will
not be tolerated. Perhaps there were instances in our evolutionary past when
a brave member of a neighboring tribe sneaked in to assess tribal strengths
and weaknesses in preparation for later warfare. Such a person would be noticed
as a “stranger” who dressed differently. A tribe whose members were tolerant
might merely shrug and leave the stranger alone, whereas a tribe with intolerant
members can be expected to challenge the stranger and demand an explanation
of who he was and what he was up to. Clearly, if tribes are in chronic conflict
conditions favor genes that predispose to intolerance. Thus, conditions
of chronic conflict should increase the incidence of two types of genes:
those that predispose to “parochial altruism” and those that predispose
to intolerance. The game theory simulation by Choi and Bowles (2007) show
that indeed both genes increase their representation in hypothetical gene
pools that are in chronic conflict.
decisively overwhelms opposing
tribes that it creates a form of peace that lasts for several generations.
The evolutionary forces that selected genes for intolerance and parochial
altruism are relaxed, and in their place are new forces that reward the opposite
genes. During peace genes are selected that predispose to tolerance and
selfishness. Again, this dynamic was demonstrated to exist in the simulations
by Choi and Bowles (2007). Wilson and Wilson (2007) as well as Turchin (2007)
have suggested this scenario as a way to understand the fate of empires. Indeed,
this is one way to view the decline and fall of civilizations.
It seems ironic
that war and peace elicit genes with opposite traits. How can these reversals
be achieved? Two modes are possible. Either the population evolves in a way
that changes the representation of “genetic types” or the individual members
take readings of an ever-evolving social setting and automatically adjust
their attitudes and behaviors. Both modes are based on gene expression, but
the latter is more sophisticated. Just as the immune system takes readings
of pathogens in the blood and adjusts its activity accordingly, the brain
is capable of reading social situations
and adjusting its activity in an adaptive manner.
There are two important
clarifications for this use of the term “adaptive.” First, something is adaptive
if it helps the genes for it to survive better. Second, the specified change
is adaptive (for the genes) provided the current setting is similar to the
“ancestral environment.”
The first clarification
conveys the message that behaviors that help genes survive may not be in
the best interests of individual welfare. Consider the switch from peace time
to war time; the individual is expected to become intolerant and hateful,
and he is expected to sacrifice his life through heroic acts that protect
the home tribe. His fellow tribesmen may benefit by this heroism, but not
the hero.
The second clarification
has become important in modern times because tribes have been replaced by
nations consisting of members from many genetic backgrounds. Japan is one of the few
nations that has preserved its genetic purity,
so there may be some genetic sense for the Japanese to engage in extreme
acts of heroism (e.g., kamikaze heroics). It is also noteworthy that the Japanese
in peace time have one of the lowest crime rates in the world. For them,
the current environment resembles the ancestral one in important respects.
But for most other nations the populations are so genetically diverse that
the genes are foolish to create individuals willing to become loyal patriots
ready to fight to the death for the Fatherland.
If humans were capable
of sanity they would mock patriotism for the pointless suffering it inflicts
upon humanity. Patriotism has always been pointless from the perspective
of the individual, but it is now also pointless from the perspective of the
group. Yet, it cannot be eradicated since it has been so crucial to genetic
survival for so many generations.
What a pathetic
situation humans find themselves in. Anyone who mocks patriotism, who points
out that it serves no purpose, will be branded “unpatriotic” – and their
message will not be heard. The need to enforce patriotism has been so strong
for our ancestors that they created a mythical entity to help enforce it:
God. This creation was instigated by the genes, of course, since they were
the beneficiaries of behaviors that secured their survival at the expense
of individuals. Since the modern “state” is an outgrowth of primitive tribes,
governed by chiefs and their helpers, it can be said that the church and
state were meant to work together. The 18th Century struggle
to separate them was motivated by a subconscious realization that individuals
were the victims of this collaboration. The separation of church and state
is a historical aberration, doomed to a short existence. Every humanist should
be sad that the few bastions of 20th Century sanity are
doomed to revert to their former evil state in the 21st.
In trying to understand
the rise and fall of empires it will be wise to keep in mind the possibility
that they are related to the rise and fall of genes that predispose for parochial
altruism and intolerance. Other factors deserve consideration. Most of the
forces causing empires and civilizations to rise and fall are based on evolutionary
changes to the genome that require an understanding of the different levels
of evolutionary selection. This chapter introduced the concept of “group
selection.” We must also consider selection at the level of the individual
and the gene. This is the goal of the next chapter.
─────────────────────────────────
CHAPTER 12
─────────────────────────────────
LEVELS OF SELECTION
AND
THE RISE AND FALL
OF CIVILIZATIONS
Abstract
Evolutionists still dispute the relative importance
of "group selection" while favoring almost exclusive selection at the level
of the gene. There is never a discussion of that in‑between level which I
shall refer to as "individual selection." This is understandable given that
individuals die, whereas genes and groups survive on evolutionary time scales.
However, I present a different
definition of "selective force" which more directly addresses the factors
influencing the fate of genes, permitting the use of the concept "individual
selection." With this modified way of viewing causes
for gene frequency changes there is a simple way to "partition" causative
factors between "levels" that I shall term Gene Selection, Individual Selection
and Group Selection (GS, IS, and GrS). The concepts GS and GrS differ somewhat
from the traditionally used meanings for gene selection (kin selection)
and group selection (multi‑level selection). I present an overview perspective
for understanding the relative importance of these three levels of influence
as they relate to the rise and fall of civilizations. I conclude that civilizations
are an anomaly that arises when individuals break loose from the most confining
bonds of the genes, as expressed by GrS, and give birth to IS. The creative
forces let loose by an era of IS propels the society embracing it to create
the thing we call a "civilization."
But history teaches us that civilizations are
short‑lived. It may be that by its very nature a society constructed upon
a base of individual freedom is vulnerable to fanatical attacks by a residual
of contemporaneous societies that remain gripped by GrS forces. This may
allow us to understand why civilizations have always collapsed, in spite
of their being surrounded by social groups with inferior levels of technology
and oppressive levels of individual subjugation. It may be theoretically
possible for civilizations to endure after the more primitive form of GrS
societies are eradicated, but human nature is such that fanatical GrS societies
will probably re‑form spontaneously. If this occurs, civilizations will be
doomed to fall after every rise, unless the long stretch of time somehow leads
to the weakening of this impulse for reverting to GrS fanaticism.
The previous chapter
got "ahead of itself" somewhat, so before proceeding further with the concept
for the rise and fall of civilizations it will be necessary to back up and
review some of the past 100,000 years and the rise of the artisan, which
set the stage for the rise of civilizations.
Introduction
In my view the sociobiology
paradigm is the 20th Century's greatest achievement.
When, for example,
a sociobiologist considers observations of Darwin's finches on the
Galapagos Islands, and the changes
they undergo in response to a year of heavy rainfall that produces foods
rewarding a different beak length, the concurrent rapid evolution of birds
having different beaks is understood using a theoretical paradigm in which
genes compete with other genes. Similarly, when prairie dogs warn of predators,
and in the process catch the attention of the predator, the altruistic act
is understood using the same sociobiology theory ("inclusive fitness") whose
mathematics was worked out by William D. Hamilton (1962). Both examples illustrate
evolutionary change as a product of competition at the level of the gene.
There is almost
a consensus among sociobiologists that any gene frequency change should be
viewed as the product of competition and selection occurring at the level
of the gene, and not higher. Genes assemble individuals as mere "vehicles"
for the genes, and these "lumbering" creations (Dawkins, 1976) compete with
each other in order to enhance the competitive prowess of the genes within.
This same theory also allows for the perspective that groups
of individuals can be viewed as "vehicles" for carrying the genes in an even
larger arena of gene competition. In other words, the accepted sociobiological
theory states that evolution occurs when gene frequencies change in response
to competition at the level of the gene, and that it is unnecessary to take
explicit account of competition at the level of the individual or the group.
Reeve (2000) seems to have shown that the equations of the "standard Hamiltonian
inclusive fitness" theory provides for group effects, thus eliminating the
need for considering groups as a level for competition.
As much as I like
this theory, and in spite of the fact that I will defend it as basically
correct for providing a proper account of essentially all evolutionary observations,
I shall consider another paradigm for "understanding" gene frequency changes.
This other paradigm has the advantage of addressing
some observations that the sociobiological paradigm is theoretically incapable
of explaining.
I assert that sociobiology's
basic task is to explain why the frequency of a gene in a gene pool changes
over time. I agree that the first order explanation must be that genes can
achieve success by creating individuals who do a better job of reproducing
those genes, and those of their kin, thus accounting for a greater representation
of these genes in future generations. However, Hamiltonian inclusive fitness
theory is an awkward tool for understanding group competition and it seems
greatly handicapped in dealing with humans who have partially "liberated"
themselves from the genetic grip by employing "logic" to influence decisions.
In the next section I present examples of cases that pose difficulties for
inclusive fitness theory, and in the sections that follow it I will suggest
a different way of viewing the locus of causation for gene frequency changes.
After presenting
these humble suggestions for amending sociobiological theory, I shall then
march forward into dangerously speculative territory, and address the recurring
puzzle of why civilizations rise and fall. In doing this I shall rely on
my newly‑defined concept for evolution at the Individual Selection level.
Special Cases That
Defy Sociobiological Explanations
Sociobiological
theory assumes that physical environments do not undergo drastic changes
on time scales shorter than can be accommodated by evolutionary adaptation.
For example, if a rare and drastic climate change occurs it is possible that
a multitude of adaptations related to earlier climates will be rendered useless
and the fate of the species (and the genes that are unique to it) cannot
be predicted. This may seem like a trivial "objection" to sociobiological
theory, but it serves to illustrate that special cases do exist for which
sociobiological theory is helpless. This illustration may not be a trivial
exception to sociobiological theory given the ever-increasing evidence in
the climate record for drastic and rapid climate changes. For example, Weiss
and Bradley (2001) list 7 drought events that led to societal collapse (10,000
BC, 6400 BC, 3100 BC, 2200 BC, 550 AD, 950 AD, 950 AD, and 1280 AD). As an
extreme example, climate can lead to the abrupt demise of species, as happened
65 million years ago when an asteroid impact created a "global winter" that
exterminated the dinosaurs and allowed mammals to flourish.
Humans present a
special case in two respects. First, they associate in super-tribes that require
strict adherence by their individual membership to arbitrary customs. The
drive for ever larger super-tribes may have been caused by the winner‑take‑all
nature of warfare that evolved sometime during the past 100,000 years. To
maintain the required superiority of numbers, and to enhance the competitive
effectiveness of large tribal groups, I suggest that the power of the group
over the individual grew to oppressive levels. An individual born into a
tribe would have no choice but to adhere to the tribal beliefs and customs
and to engage in coordinated warfare with neighboring tribes. The tribes with
cultures that evoked a high degree of fanatical loyalty to tribal endeavors
would be more successful at surviving and dominating their region. In this
setting the individual (and his genes) experience a high degree of "shared
fate." The group and its membership would prevail or perish together.
In this setting
a novel genetic mutation that began by affecting just one person would be
rewarded far less than in a setting where group‑imposed behaviors were weaker,
or not present. If we ask "what factors affect the fate of genes in the setting
where fanatical tribes are in constant conflict, where there is an ever-present
risk of the entire tribe’s extermination?" we are forced to answer that
"the perspective of selection at the level of the group appears to be more
useful than selection at the level of the gene." In other words, when individuals
are severely subjugated by the imperatives of tribal survival we must reckon
with more than just genetic mutation in order to conveniently account for
changes in gene frequency over time.
Humans confound
sociobiological theory in yet another respect. With the evolution of a "logic
using" left cerebral hemisphere, or left brain (LB), some individuals have
achieved a modest level of liberation from the influence of the genes. "Rational"
decisions are to a large extent "genetically unanticipated," which in some
small measure disconnects the fate of the genes, for which the individual
is a vehicle, from the ancestral environment selective forces that guided
the development toward this wondrous, rational LB. One dramatic and straightforward
example is the decision by an individual in contemporary society to use birth
control measures to limit reproduction. When the genes created smarter brains
they had no way of anticipating that those brains would subvert the genetic
agenda. The sociobiological literature is inexplicably quiet on this confounding
factor.
Before presenting
a new way of viewing "selective forces" that incorporate the two anomalous
aspects of human evolution described above I want to present a brief history
of the changes in thought on where the locus of power for genetic change
resides. There has been an active debate over the causes for gene frequency
changes, and there have been several shifts in the preferred way of assigning
importance to the various levels at which selection can occur.
Brief History of
Level of Selection Viewpoints
Before Darwin's 1859 book, On the Origin of Species, most people believed that God made
humans in such a way that our behavior guaranteed the survival of the species.
Darwin displaced God from
this thrown, and groped to identify what replaced Him. The concept of genes
was too vague in Darwin's time to be incorporated
by his theory, since Gregor Mendel's article "Experiments With Plant Hybrids" (1866) lay unopened on Darwin's shelf; the rest
of the intellectual community also failed to appreciate this work until 1900,
18 years after Darwin's death. For this
reason Darwin can be excused
for writing in 1859 "natural selection works by and for the good of each
being." Nevertheless, with this statement Darwin appears to place
the locus of influence at the level of the individual. Later, Darwin shifted toward
group selection when he wrote (1871) "primeval man regarded actions as good
or bad, solely as they obviously affected the welfare of the tribe, not of
the species." Alfred Russell Wallace, who co‑discovered natural selection
as an explanation for evolution of species, "stressed that group selection
... played an important role..." (see Merlotti,
1986). Darwin, it seems, eventually
joined Wallace in giving group selection a leading role in natural selection.
According to Carl
Sagan and Ann Druyen (1992, p. 70), "One of Huxley's interests had been the
idea that all animals, including us, were 'automata,' carbon‑based robots,
whose 'states of consciousness... are immediately caused by molecular changes
of the brain‑substance.' Darwin closed his last
letter to him with the words: 'Once again, accept
my cordial thanks, my dear old friend. I wish to God there were more automata
in the world like you.'" (see also Huxley, 1874).
This idea was "ahead of its time" and did not become part of the climate
of opinion in the late 19th Century. However, I hope this "reductionist"
idea will be resurrected during the 21st Century and will form the basis
for understanding all behavior.
By 1950 several
writers rediscovered the importance for natural selection of inter‑group
competition and, hence, intra‑group cooperation. Merlotti (1986) summarizes
Spencer (1892) as believing "Let enough members of a society disobey the code
of amity (for members within the tribe) and the society will fragment; let
enough disobey the code of enmity (against neighboring tribes) and the society
will be crushed." Merlotti quotes Sumner (1906) "The exigencies of war with
outsiders are what makes peace inside, lest internal discord should weaken
the we‑group." Sir Arthur Keith wrote persuasively in the same vein (1946,
1948), as again summarized by Merlotti "the success of the human species had
been secured by cooperation within groups and competition between them."
The following table
will be useful in seeing how differently thinkers of different periods partitioned
the locus of influence for natural selection.
TABLE
I
%
Importance
|
<1859 1859 1950 1962 1966
1994 Present
|
|
GOD
100 0 0 0 0 0 0
|
|
SPECIES 0 0 0 80 0 0 0
|
|
GROUP 0 20
80 15 10 20 10
|
|
INDIVIDUAL 0 80 20
0 0 0 10
|
|
GENE n/a ?
? 5 90 80 80
|
By the mid-20th
Century the forces of academic Marxist influence began to take their toll
on the quality of anthropology thought. Whereas most previous theory incorporated
an inherited predisposition to affiliate with tribes, and to be predisposed
from birth to adhere to Spencer’s "tribal mentality" (i.e.,
with amity toward fellow tribesmen and enmity to all others), the climate
of opinion shifted during the 20th Century toward a form of "cultural
determinism." Biology was "out" and culture was "in." Culture was seen as
a guarantee of species survival, and the locus of influence was "whatever
is good for the species." The good of the species was such an appealing thought
that those who could not relinquish a role for a "species nature" tried
to see a pattern of evidence that instincts served species survival goals.
Perhaps the most comprehensive expression of this idea is the 1962 book
Animal Dispersion in Relation to Social Behavior by Wynne‑Edwards. It amassed a tremendous amount of data in support of the
idea that when a species begins to over‑exploit its environment individuals
will reduce their rate of reproduction (voluntarily) as if motivated to
guarantee that resources will be available for future generations.
The fundamental
flaw in this idea is that organisms are gene‑created automata, and they cannot
perceive the future, or even care about the future; they behave the way
the genes have programmed them to behave, reacting to environments in ways
that are programmed, and the genes that constructed them are the ones that
have been the most successful proliferators in past gene‑upon‑gene
competitions. Culturgens (culture’s "memes") are also a factor for human
behavior, but even memes cannot be credited with caring about the fate of
the species.
Robert Ardrey's
writings have withstood the test of time, in spite of the ridicule heaped
upon him by those who resented his audacity for having an opinion on anthropology
after having established himself in a different field (playwright). From
his 1961 African Gensis to his 1976 The Hunting
Hypothesis, the amity‑enmity duality was a central theme from which he
argued that aggression is a natural human instinct, as were within‑group cooperation
and loyalty. He wrote "If competition takes place not only between individuals
but between groups, then the group with greater endowments of loyalty, cooperation,
self‑sacrifice and altruism concerning social partners will be selection's
survivor." Thus, Ardrey tried to keep group selection "alive" during the
1960s and 1970s, but due to his background as a playwright he was not taken
seriously by the Marxist anthropologists of that time.
During the first
half of the 20th Century the discipline of physics made dramatic
advances that captured the imagination of the general public. The climate
of thought by mid‑century should have been congenial to the notion that physical
events at the atomic level dictate all particle motion, and therefore all
animal behavior. There is no evidence (that I am aware of) that people were
thinking this way by mid‑century, even though Huxley had suggested the idea
80 years earlier. With the discovery in 1953 that genes are double‑helix
DNA molecules the stage was finally set for thinking of gene‑assembled organisms
as automata. The slowness of the process by which humans approach Truth is
best understood by remembering that every creature is a gene‑created automaton,
and that a human is programmed to think in ways that served the genes that
constructed his ancestors. Therefore, if the comprehension of a fundamental
truth has never influenced the selection of human ancestors we cannot expect
that humans will quickly grasp that truth. The gene‑centered perspective
requires a difficult leap of imagination for which the human ancestral environment
has not prepared us.
William D. Hamilton
(1964a,b) was one of the first writers to grasp
that evolution should be viewed as occurring at the level of the genes! He
created the mathematical foundation for understanding how evolutionary competition
at the genetic level can explain such social behaviors as altruism. Hamilton's "intrinsic fitness"
theory is a “mechanistic” theory for the evolution of individuals by natural
selection that behave "altruistically" toward each other, provided the interactants
are close relatives.
The math of Hamilton's derivation is
daunting, even for mathematicians, and fortunately George C. Williams came
forward with a non‑mathematical interpretation of Hamiltons' message (plus
other implications of the gene‑centered view) in a landmark book Adaptation and Natural Selection (1966). Williams addressed
the issue of levels of natural selection, and allowed for the theoretical
possibility that group selection, GrS, could occur when certain conditions
existed. He considered conditions that must exist before GrS can occur (my
description of this is adapted from Buss, 1999).
According to Williams,
for group selection to be important there must be:
1) a high degree of "shared fate" of the members of the
group,
2) low levels of reproductive competition within the group,
and
3) recurrent patterns of differential growth and extinction
of groups.
Williams demolished
the old version of group selection, wherein adaptations evolved for the benefit
of the species. He was even skeptical that this new version of group selection
could be found in nature, for he believed that the three conditions he specified
were rarely met, especially with humans.
Edward O. Wilson's
book Sociobiology: The New Synthesis
(1975) and Richard Dawkins' popular book The Selfish Gene
(1976) consolidated the perspectives presented earlier by W. D. Hamilton,
G. C. Williams and their lesser‑known predecessors (R. A. Fisher, J. B. S.
Haldane, S. Wright). The notion that natural selection worked at the level
of the genes, and not groups, grew in strength with their writings.
Wilson and Sober
(1994) revived group selection theory, and emphasized that groups can be
portrayed as "vehicles" for genes, somewhat similar to the way individuals
are vehicles for genes. They did not challenge that selection at the level
of the genes is important; rather, for social animals it is also necessary
to incorporate the effects of groups in order to understand the fate of the
genes carried by its members. This version of group selection theory is sometimes
referred to as "multilevel selection theory" (MLS). If
the idea of multiple levels for viewing selection bothers you, re-read Chapter
1, and Appendix A, where I argue that a proper understanding of reductionism
allows for more than one “level of physical explanation.”
Even though MLS
embraces such levels as the gene, the group, and even the species (and multi-species
ecosystems), it excludes the individual. A 1998 book by Sober and D. S. Wilson,
Unto Others: The Evolution of Unselfish
Behavior, endeavors to place group selection on a sound, mathematical
footing. Although Sober and Wilson acknowledge that
the mathematics of Unto Others is "equivalent" to the mathematics
of Hamilton's inclusive fitness
theory, they claim that the rearrangement of terms in the equations renders
it a better tool for understanding the role of competition at the group level.
Reeve (2000) argues that the equivalence of the two mathematical formulations
renders the group selection arguments in Unto Others unnecessary,
since both theories are based on competition that ultimately occurs at the
level of the genes. The latest version of "group theory" thus portrays the
argument as a subjective preference for the mathematical arrangement of
terms in an equation.
Overview of Human
Evolution
I accept the conventional
wisdom that before approximately 125,000 years ago humans lived in tribes
of approximately 50 to 150 individuals. Inter‑tribal conflicts may have been
common, but I am unaware of compelling evidence that tribes exterminated
each other before approximately 40,000 years ago (Keeley, 1996). It is not
known if individuals were free to switch tribes. The simplest model for human
evolution in those times would be to place the bulk of selection pressure
at the level of the genes. To the extent that primitive humans lacked "culture,"
their evolution and the evolution of all other animal and plant life can be
explained just as successfully using gene‑level sociobiological theory.
At some time between
approximately 70,000 and 12,000 years ago human tribes began to grow in size
dramatically, and most humans soon found themselves members of "super-tribes"
several times larger than the original tribes. The drive toward larger tribes
was irresistible when they became more effective in inter‑tribal warfare.
The super-tribes also could more easily afford to allow a small number of
men to specialize in full‑time weapon and toolmaking, and other artisan specialties.
Whenever inter‑tribal warfare began to result in the extermination of the
losing tribe, sometime between 70,000 to 12,000 years ago, that was the
time when tribal size and tribal loyalty became crucial determinants of
the fate of genes. Since the individuals within a tribe had a "shared fate"
when tribal decimations began we can begin to consider placing the rise
of "group selection" (the modern version of GrS) to this critical time. We
may also speculate that tribal rules would grow more restrictive at the same
time. Genes for "compliance" would be rewarded, for they would produce individuals
who could embrace conformity with a minimum of cognitive dissonance (Lumsden
and Wilson, 1981, and Boyd and Richerson 1985).
Super-tribes, and
the growth of a tribal culture requiring strict adherence, represent a critical
stage for the shift of evolutionary relevance from the genes to the group.
Fanaticism, reinforced by tribal custom and religious fervor, reward the
genes of individuals within that group, for they make the tribe a more formidable
enemy to their neighbors (Kriegman and Kriegman, 1997). Individuals within
super-tribes have no viable alternative to membership in their tribal group.
After this transition, occurring sometime between 70,000 and 12,000 years
ago, the factors that determined the fate of genes would be shared between
the level of genes and the group. Prior to this transition most of the factors
determining the fate of genes would have been at the level of the genes.
From the time of
the transition to super-tribal conflicts to the time of Classical Greece,
the factors determining the fate of genes would have been shared between
the level of the genes and the level of the group, between GS and GrS. The
notion of individual rights, or individual liberation from the oppressions
of tribal life, began during the 6th Century BC in the area of northern Greece. It might have
had beginnings in the Minoan civilization, but we have no records of what
people believed from that far back (ending abruptly when the volcanic island of Thera exploded in 1628
BC). After the slow collapse of the Roman Empire the rights of the
individual lost influence and group rule was restored. With the re‑discovery
of Greek philosophy by the 17th Century Enlightenment philosophes
it again became fashionable for the individual to assert himself. The church's
power was at an all‑time low when this 17th Century rebirth of individualism
occurred. What is now referred to as Western Civilization is an outgrowth
of the ideas originally expressed by the Greek philosophers.
Western societies
at the end of the 20th Century gave individuals the freedom to
move about, to experiment with alternative cultures, and to think thoughts
that contradicted those generally accepted. Tolerance of individual differences
and respect for individual freedoms were at an all‑time high at this time
in the Western world. For example, a person was free to adopt birth control
measures to control pregnancy, and many educated and wealthy individuals
choose to forego having offspring. The fate of the genes was influenced by
individuals who were free to make rational decisions that were often dictated
by a desire to optimize the welfare of the individual at the expense of genetic
propagation. This short‑changing of the genes was accompanied by a weakening
of the group to which the liberated individuals loosely belong, as will be
explained below.
A New Measure for
the Strength of Selective Forces
This section may
be "tedious" for most readers. I will summarize it in the next paragraph
so that you may skip to the next section whenever you think you're encountering
more detail than you need.
The task of this
section is to assign importance to each of three levels for selection (gene,
individual and group) in an account of why genes change their frequency in
a gene pool over time. If the gene level is all important, then we will end
up stating that: the "force of the genetic level"
FG = 1.00, the "force at the individual level" FI = 0.00,
and the "force at the group level" FGr = 0.00. However, if group selection
is important, there will be a re-partitioning of selection strength so that
genes will be a more important explanation for gene frequency changes; for
example, we would state that FG = 0.50, FI = 0.0, and FGr = 0.5. Note that
the sum of the three forces adds to 1.00, which is a condition I impose for
convenience. The task is to "partition importance" among the three candidate
levels in order to explain what is causing a gene pool to evolve.
In groping to define
a new "measure" for use in explaining changes in gene frequency let us review
some of the attributes we expect of it. If this resembles describing the
answer, then formulating the solution, it is. Common sense should sometimes
guide us.
Consider the situation
before there were super-tribes that held individual members captive by inhibiting
the full measure of unique individual attributes and inhibiting the free
expression of personal aspirations. It should be clear that however this new
measure is defined it should assign an almost complete amount of strength
to the level of the gene and very little to the group. After super-tribes
came into existence, we want this measure to share strength between the level
of the genes and the level of groups. After individuals begin to liberate
themselves from group‑conforming societal pressures, a phenomenon which has
occurred most dramatically in Western cultures, we want this measure to assign
some strength to the level of the individual. The individual who is free to
choose his culture, his role in that culture, and his reproductive lifestyle,
warrants a seat at the table of power over gene frequency changes.
The measure I propose
is based on asking "What factors affect the fate of the genes? And how are
these factors partitioned between the level of the gene, the individual and
the group?" If we constrain the three strength values to add up to 1.0, then
as individual liberation grows, for example, the strength of the genes and
the group diminish. Thus, FG + GI + FGr = 1.00,
where FG is proportional to the sum of selective forces identified as originating
at the Level of the Gene, FI is proportional to the sum of selective forces
identified as originating at the Level of the Individual, FGr is proportional to the
sum of selective forces identified as originating at the Level of the Group,
and the normalizing of the three forces is done so that the sum of forces
= 1.
In this treatment
I ignore all factors that influence the fate of genes that do not belong
to the above three categories. For example, I will exclude from consideration
earthquakes, floods, drastic climate changes, asteroid impacts, and all other
rare physical environment events, even though they do indeed occasionally
affect gene frequencies. By neglecting them in this analysis I am following
the precedent of most other evolutionary models, and my treatment merely
fails to capture random perturbations which will not affect the conclusions
I wish to make. It would be straightforward to formulate a version of this
theoretical treatment that includes these environmental catastrophes, but
it is not my intention to include them here in order to better emphasize the
role of the non‑gene levels of selection, GrS and IS.
The method for measuring
"Selective force," which I shall describe momentarily, is inherently subjective,
and this is the weakest part of my argument. However, it has attributes that
make it useful for things that other formalisms do not allow. At any given
time many things are happening that may influence subsequent changes in gene
frequency in a gene pool. The basic task is to associate changes of gene
frequency with causes for those changes. Instead of attempting to ascribe
cause and effect through an explicit treatment of each gene's phenotypic
expression, and speculating about the implications of that altered phenotype,
I propose to employ mathematical tools that are blind to the mechanisms for
cause and effect. I propose to perform a multiple regression analysis of
all gene frequency changes in a genome, over a specified time, using as independent
variables all parameters that can be measured and that describe potentially
relevant aspects of the social environment, the genes found in individuals,
the milieu of culturgens that individuals are exposed to, "novel thoughts"
experienced by individuals, and many other similar properties that could
in theory be measured. This item "novel thoughts" may be troubling, but I
want to retain it for reasons that will become clear later.
Obviously it is
not feasible for anyone to measure all relevant parameters describing the
social environment, the genotype of individuals, the cultural milieu, and
the thoughts experienced by individuals, but let us suppose for the sake
of argument that these parameters nevertheless exist. I make this request
of the reader in the same spirit that is required by twin study investigators,
for example, who attempt to partition the effects upon individual traits
by genetic versus environmental causes. In those studies it is not necessary
to identify every factor that influences how a person becomes who they are;
rather, it is merely assumed that a myriad of such factors in each category
exist, and the investigator proceeds to partition causation of the aggregate
of effects by performing correlation analyses. The following paragraphs are
the ones the casual reader may wish to skip.
Assume that we create
an immense inventory of parameters that describe the state of a "setting"
in space and time. Assume further that we assign each parameter to the categories
G, I, Gr and "other," and
reject all parameters belonging to "other," such as natural catastrophes
(for the reasons presented above). The number of parameters belonging to G,
I and Gr shall be referred to as N. The "state" at any given time is an N‑dimensional
"state vector," to use mathematical terminology. After choosing a "timescale"
for associating the state vector with events of a gene frequency change we
can, in theory, perform a multiple regression analysis (MRA) for each gene.
Each MRA will use gene frequency as the "dependent variable" and the N‑dimensional
state vector as elements for N "independent variables." Each MRA will then
produce N correlation coefficients, one for each parameter. We now can sum
the correlation coefficients in the following way:
FG' = Sum of all correlation coefficients associated with
Gene Level parameters,
FI’ = Sum of correlation
coefficients associated with Individual Level parameters,
FGr'=Sum of all correlation coefficients associated with Group Level
parameters.
which are the un‑normalized
"forces," that are easily normalized by dividing by their sum.
The entire procedure
just described is then re‑performed for the next gene, and so forth until
all genes are thusly treated. Since only a few dozen gene loci have more
than one allele (for a given species), this analysis need only be repeated
for a few dozen genes. These several dozen results are combined to arrive
at an overall score for the relative importance of each of the three levels
of selection.
Again, I want to
emphasize that it is not necessary for the above procedure to be feasible
in order to make use of the concepts that they convey. I only ask that you
accept that it is conceptually feasible! In some sense I
am appealing for a belief in concepts that are just as "real" as the coefficients
in Hamilton's horrendously
complex mathematical derivations. If we can imagine specific coefficients
to exist in a hypothetical world, then they do exist in the real world even
though as a practical matter we are limited to only an approximate measure
of them.
One further clarification
is needed here: there is nothing in my proposed paradigm that is incompatible
with present‑day sociobiology theory. What I am suggesting is an alternative
way of viewing events. This is often done in physics. For example, a physical
chemist can, in one situation, treat a salt crystal as a lattice structure
held together by electrical forces, while in another
situation treat the same salt crystal as a group of sodium and chlorine atoms
that can become dislodged for chemical reactions when dissolved in water.
The physical chemist knows, during both treatments of the salt crystal, that
whatever happens is the result of the four forces of nature acting upon
tiny masses, in a way that is too cumbersome for practical use in everyday
experiments (the four forces of nature being gravitational, electromagnetic,
weak and nuclear). Similarly, the person trying to understand human behavior,
or the rise and fall of civilizations should know that every person's actions
are dictated by the same four forces of nature acting upon tiny masses. I
acknowledge the frequent need to seemingly overlook the inherently reductionist
nature of all phenomena in order to advance our "understanding" of the everyday
world. Sociobiology and my suggested partitioning of influence among three
levels is just another example of looking at the same phenomena from different
perspectives, and they are NOT contradictory.
Levels of Selection
and the Rise and Fall of Civilizations
Consider the following
figure, where I have marked off 9 stages, "A" through "I", that I am suggesting
typify the evolution of a human civilization.
Figure 11.01. Hypothetical
allocation of "selection strength" for the three levels Genes, Individual
and Group.
At stage "A" we
are to imagine that mostly the gene level of selection is important in determining
the fate of genes. At this stage the individual does not assert himself,
he does not make birth control decisions, or decide to walk away from the
tribe and live alone. Also at this stage there are no super-tribes, and tribal
conflicts are not all‑or‑nothing group exterminations. Consequently, the strength
of group selection pressures is very small. It is only non‑zero because I
assume that the fate of individual social alliances has some effect upon
the survival of the individual and his reproductive outcome.
At stage "B" we
have super-tribes exterminating each other, rewarding the super-tribes that
enforce conformity among its membership. The fate of the genes within an
individual are less affected by the individual phenotypic expression of them,
for some of the individual's destiny is beyond his control by virtue of the
fact that he belongs to a tribe that will survive or be extinguished on the
basis of how well the tribal membership works together. It will matter only
slightly that a particular individual is greatly endowed by his genes if the
tribe he belongs to is ineffective in combat with its rivals.
By stage "C" the
group has become oppressive in its restriction of individual members. Any
deviation from the cultural norm will be punished, so any differences in
individual genetic profile, any outstanding abilities for example, are ignored
and each individual is subservient to the dictates of group needs. This stage
is marked by devotion to tribal rituals, unquestioned loyalty, fervent religious
devotion, fanatical fighting and a readiness to sacrifice the self for the
greater glory of the group. For a modern Westerner this stage is the most
difficult to like. Kriegman and Kriegman (1997) suggest that religion was
an invention that enhanced the fighting competitiveness of a group because
it provided a "rationale" for fanatical behaviors; any groups not having a
religion to motivate fanatical adherence to the group's destiny would be handicapped
during warfare. I incorporate this thought as a crucial component of my argument
that civilizations are destined to be short lived in a world where fanatical
societies exist.
Stage "C" represents
the "birth of individualism." It is no coincidence that the Olympic games, which emphasize individual as opposed to team
competition, originated in a region that gave birth to the notion of celebrating
the individual. The Greek philosophers discussed the proper relationship
between the individual and society, and the proper role of a government. Democracy
as a form of government is an outgrowth of a shifting of power from a "tribal
leader with group support" to the individual. When the individual is set
free to achieve, and receive credit for his achievements, it should be no
surprise that more achievements per capita should result. Commerce and technology
should develop faster, and more economic niches should be created. In a society
where the individual has government sanctioned rights, as in a democracy
and free‑enterprise economy, there should develop a greater tolerance for
people having new ideas. Productivity should rise in not only the commercial
sector of the society, but also the intellectual. Literature, the arts, and
philosophy are individual endeavors that attest to a vibrant social order
that rewards individual initiative. These are the conditions that lead to
what we call a "civilization."
But as the individual
thinks for himself, he exerts an influence over gene frequency changes as
well as the shape of society, and by the zero‑sum nature of my proposed partitioning
of the forces of selection we must see a decline in the influence at the
level of either the genes or the group. I suggest that the group is the big
loser, and perhaps its losses are so great that even the genes are winners.
During the rise of a civilization, when more power flows to the individual,
the genes can still be winners because an individual with a new mutation
has the potential for prospering more than the average of his society. Even
today the extremes of personal wealth continue to widen. Bill Gates has more
wealth than 1 million average people in the Western World, and his wealth
is based on genetic intelligence and business savvy (and, yes, the luck of
an opportunist); at least you will agree that it is not based upon inheritance
or acquisition through plunder.
It should be pointed
out that Stage "C" appears to be driven by a bold assertion of a growing
minority of people with "strong" left brains! Any neuropsychologist would
agree that the manner of discourse exhibited by Socrates, Plato and most other
philosophers of that time can be explained by invoking a leading role for
styles of thought that only the left cerebral cortex is capable of performing.
Elsewhere I have written that during the past 70,000 years, at least, there
was a growing place for artisans within tribes, and these artisans could perform
their work most effectively if they had well‑functioning left brains. I have
also argued that the left brain, as well as the frontal
lobes, have undergone the greatest amount of evolutionary change in
recent times, and that this is due to the need for full‑time artisans for
the support of tribal endeavors, and that this has grown in importance during
the past 70,000 years.
Stage "D" is a growth
of what was started in Stage "C." During this stage individual liberation,
made possible by the influence of people with left brain styles of thinking,
enjoys a toleration from society at large that is unprecedented. Tolerance
for new ideas, new customs, and challenges to old ideas and customs becomes
acceptable and expected. The lifting of group oppressions allows a release
of unprecedented creativity and productivity, and this energizes commerce,
technology and government efficiency, which leads to a spread of greater
material wealth throughout most of the population. Excesses of wealth, found
in both successful individuals and a bountiful government, benefit the arts.
Stage "D" meets the dictionary definition for civilization, as "An advanced
state of intellectual, cultural, and material development in human society,
marked by progress in the arts and sciences, the extensive use of writing,
and the appearance of complex political and social institutions."
But a curious thing
happens during the progress toward a more extreme development of Stage "D"
civilization, as we are now experiencing in Western Civilization. The recipient
of civilization's bounty, the individual, turns inward, and becomes absorbed
with personal, individual well-being. Beyond the boundaries of civilization's
campfire exist uncivilized societies that have
not absorbed the values of their more successful cousins. These societies
are on the fringes of the fountain of wealth, and they feel "used" and left
behind, as they pick up the crumbs that fall their way. Instead of wanting
to emulate those better off, they resent them, and they wish to defeat their
well‑off neighbors, and perhaps plunder the fruits of other men's labors.
They are moved by the ancient and primitive tradition of seizing what one
wants instead of producing it.
While resentment
grows among those relegated to being spectators of the civilized, and while
the numbers of those lucky civilized members grow, another unexpected force
gathers strength from within the civilization: it is a curious cadre of "cultural
enforcers." These people are a residual of past episodes of boom and bust,
and their ancestors have saved their kin from the excesses of success. The
cultural enforcers (religious fundamentalists) wish to curb the undisciplined
pursuit of civilization's glitter by re‑instituting some old fashioned values.
They can be likened to a well‑meaning friend trying to sober‑up a hung‑over
celebrant, as if the celebrant was merely preparing himself for the next
day's battle. For there is always a next day's battle, and every civilization
must be ready for it, or perish.
Stage "E" is the
turn‑around, a reckoning with the consequences of success. The cultural forces
from within are creating a "group mentality" in readiness for battle, and
the forces beyond civilized borders are probing their enemy for weaknesses.
Both sides silently gather strength, like the quiet before a storm.
The Stage "F" collapse
will be faster than the rise, for destruction is always easier than production.
Fanatics chip away at the existing structure by attacking places that are
most vulnerable; but just as important, they inexorably reduce public confidence
in the existing order. For those at ground level, measuring change by moments
of a lifetime, the changes may not be apparent and their significance will
not be appreciated. Someone still drives the trucks, repairs the streets,
and constructs the houses, though it is a different person and he worships
differently. Traditional battle lines will not exist, as the war is one of
skirmishes by well‑organized, small groups of fanatics, as they erode a
structure built by processes that are quietly disappearing. Instead of energies
being focused on new and more glorious projects for the future, energies
are focused on repairing crumbling social structures that directly impact
personal well‑being and on protecting society from random terrorist attacks.
The "vision" of future things is replaced by a need to make concrete repairs
and protect the security that was taken for granted in the past. There is
no time or energy for the arts, for music, or new ideas. As civilization dissolves
and eventually evaporates, it leaves a residue of useless scum.
Few people will
recognize their loss as a loss, for by then most people will have turned
over their left brains to the control of their rights. Part of the war effort,
waged by the attackers and the attacked, is a covert campaign to discredit
left brain styles of thought and left brain values. The attackers do it because
they've never known the left brain's ways. The attacked do it because priorities
now require that everyone become engaged in only essential endeavors. The
essential endeavor is defense, and defense is most effective when the postures
of fanaticism are adopted. And the retransformation to fanatic postures
requires that the left brain style of thinking be abandoned.
Stage "G" is a complete
deliverance of the once victorious civilization to the leaders of the group
mentality. The tolerance for new ideas is lower than at any previous stage,
and the individual expression of anything new and potentially upsetting to
the grip of the group is unthinkable. Religion's job is to enforce this policy,
to keep individual thoughts suppressed in order to preserve the status quo.
The individual is not the only loser, for the fate of genes is even more
strongly influenced by the vagaries of group culture. Whatever preserves group
survival and dominance defines the way things are. Genetic mutations that
in prior times would have brought their lucky individual vehicle to a winning
place, thanks to their individual creations being inclined to be creative
or productive, now produce individuals who are burdened by their superior
creativity and individual passions. The newly strengthened group selection
forces inhibit what we now call progress, for groups reward things that lead
to Dark Ages. Where Group Memes Rule, the genes are in repose, and the individual
is in eclipse.
Stage "H" and its
eventual deliverance of its victims to stage "I" (equivalent to Stage "A")
is a slow process, much slower than indicated by the figure. During this
stage there is a lessening "need" for oppressing those who threaten stability
by wanting to be progressive. The need to suppress individuality at this late
stage is less than before because few people remain who remember how to assert
one's individuality. Individualism during the Dark Ages is publicly non‑existent.
It may exist, but only furtively. Individuals who show more initiative, and
who are motivated to achieve, slowly infiltrate the positions of power, and
through a neglect of enforcing the culture of oppression they allow increments
of change to erode the power of group culture. Perhaps we are lucky that
it only took one millennium for the Dark Ages that followed the fall of the
Roman Empire to give way to
a rebirth of individualism. Part of the credit for the weakening of group
culture goes to the plague of the Black Death, which so dispirited the populace,
and so undermined the credibility of religion, that an opening appeared for
individual voices to speak out in favor of ideals that had not been publicly
uttered for 1000 years (Cartwright and Biddiss, 1972).
The longer Stage
"H" endures, the more likely it is to be replaced by changes brought about
by the voice of individuals. Evolution has produced brains that will not
stay quiet forever, and this restless energy will break through religion's
oppressive "blanket" eventually. A millennium or more may be required for
this recovery.
The current episode
of civilization shows signs of decline, amidst isolated surges of forward
growth. If the Western World's civilization reached its peak in the early
20th Century (see Chapter 15), then the complete cycle, from Stage
"A" to its repeat as Stage "H," requires approximately 2500 years. The previous
cycle may have been interrupted by the volcanic eruption of Thera, destroying
the Minoan civilization in 1628 BC ‑ which had many of the features of the
later Greek civilization. If the Minoan civilization had unfolded naturally,
and had undergone a decline caused by a human restoration of group culture
enforced by religious oppression, then the human spirit might have lain
fallow for longer than the 1000 years that in fact was required for the
resurgence of an individual‑based culture, as occurred with the Golden Age
of Greece. Thus, our present knowledge of the human record denies us the
opportunity of knowing whether a 2500 year cycle is typical. Most of the
time of our most recent cycle was spent in the Dark Ages mode. We do not
know if this was true of the previous cycle, or if it will be true of any
future cycles.
The dynamic just
described occurs at lesser levels when regions are isolated for long periods.
Thus, there are other examples to learn from of the exchange of power between
the levels of the group and the individual (always at the expense of the
gene). Asian history might be revealing in this regard.
Oscillations as
a Transitional Mode
Before 11,600 years
ago, when the transition to our present interglacial was complete, the oscillations
between Group and Individual power, each borrowing from Gene power, probably
did not exist. It is a property of some physical systems that they undergo
a transitional mode of oscillation during their shift from one mode to another.
Before the Holocene (i.e., our present interglacial), human
societies were probably exclusively tribal, and super-tribal, corresponding
to Stage "A" in the above figure. It is natural to ask "Will humans some
day be exclusively civilized, remaining so for long periods?"
For this to happen,
according to the ideas of this chapter, it would be necessary for a civilization
to "include" the entire world's people in the benefits of being civilized.
Such a condition would remove one of the energizing motives for fanatical
attacks upon civilization's structure. To be sure, some people can be counted
upon to hate the established order, no matter how beneficial it is to its
individual membership. But there must be a critical amount of discontent
for it to become a serious threat to the majority. This should be the hope
of every civilization, that it can share the benefits broadly enough that
the number of malcontents will not find each other in sufficient numbers to
constitute a serious threat to maintaining the civilization.
Alternatively, those
wishing for the longevity of civilizations may hope that the genes for malcontent
will diminish during a civilization's existence. Clearly, it is too much
to hope that a civilization will pro‑actively alter the genetic composition
of its citizenry. Unless, that is, the civilization is small in numbers,
serious about survival, and physically isolated from its neighbors ‑ as might
occur some day when settlements exist in space.
If civilizations
are to avoid falling soon after rising, they must confront both challenges:
1) attack from uncivilized, group culture societies that feel threatened
by civilization's presence, and 2) the ever‑present threat of indigenous malcontents
coming together to form fanatical cadres bent upon destroying the civilization
from within. I take an agnostic stance on the likelihood of either condition
being met someday. The human species is an experiment, and it’s not over
yet!
Acknowledgement
For This Chapter
I want to thank
Dr. Daniel Kriegman and Orion Kriegman for letting me read their unpublished
manuscript that expands upon their 1997 HBES Conference presentation. My
thinking was helped by their idea that religion's proclivity for producing
fanatic proselytizers and defenders gives religious societies an advantage
in prevailing over neighboring societies with non‑religious beliefs.
APPENDIX TO THIS
CHAPTER
A Fuller Explanation
of Group Selection
How might GrS be
measured? In the body of this essay I presented a conceptual version of a
multiple regression analysis procedure for measuring the "force of group
selection," FGr. That derivation was meant to illustrate a concept, and not
meant for use in any specific situation. However, it may be possible to crudely
measure FGr at a specific location and time. Imagine the existence of a questionnaire
with weighted scores that probe key aspects of the setting in question. Although
I leave the task of creating such a questionnaire to someone else, I shall
hint at it with the following examples.
1) How often do individuals suffer from their affiliation with a specific
group ("shared fate")?
2) How often do individuals publicly question beliefs that are held
by the majority of group members ("compliance")?
3) Are individuals free to change group affiliation without sanction;
may they have more than one group affiliation
at the same time ("membership enforcement")?
4) Is an individual free to leave the group and live without any group
affiliation (relating to the threat of a group to punish individuals by
"banishment")?
Scoring societies
using such a test might be useful in studying the "civilization's evolution."
If Western Civilization can postpone that “rounding the corner” from Stage
E to Stage F during the 21st Century, such studies might actually
be conducted.
A Fuller Explanation
of Individual Selection
It’s easy to make
the case that walking sticks and spiders that exhibit "male sacrifice" during
procreation are enslaved to their genes (male sacrifice is when the female
literally begins eating the male's head and other body parts after copulation
has begun, partly to better nourish the development of eggs but also to assure
that other males will not fertilize the female’s eggs). With humans, the
case is more difficult to make, but an earnest effort will be repaid. At
the present time the sociobiological literature merely hints at this fact,
for the field could lose public funds if it pushes forward too fast. Let us
be bold, and accept the notion that humans in all societies are to some extent
"used" by their genes, that emotional payoffs are meant to encourage individual
performance of the most essential if not dangerous, laborious and illogical
of tasks needed by our genes for gene survival. This book issues a "call
to arms" for individuals to liberate themselves from genetic enslavement.
Imagine that another book exists that calls
for the individual to also free himself from the grip of "the group" (using
whatever definition one likes for "group"). For now let us just assume that
the individual is to some extent in the grip of both the genes and the group.
In what ways are individuals now liberating themselves from these twin enslavements?
An individual asserts
his "rights" when, for example, birth control measures are used. An individual asserts
his rights when he argues for peace over war, and avoids being drafted into
non‑defensive, aggressive wars. An individual aspires to liberation from
the genes when he thinks critically of conventional beliefs and pursues thoughts
freely ‑ as a "free thinker" does. These few examples reveal the possibilities
for individual liberation from the grip of the genes and the group.
It is a novel situation
when individuals achieve some degree of liberation from their genetic and
group enslavements, and freely make personal decisions that can affect the
fate of the genes for which they have become "newly uncooperative" vehicles.
Because such individuals are less likely to produce their quota of offspring,
or to nurture their nieces and nephews, these individuals would be viewed
by the genes, if they had a view, as "freeloaders." They are not paying the
price for admission to Life!
To the extent that
other individuals remain enslaved, and to the extent that the inclination
to liberate oneself is influenced by the individual's genetic makeup, the
existence of liberated individuals will alter the fate of genes. The present
time, as we approach the Stage "E" crest of Western Civilization, is an ongoing
experiment. Let us hope that insulated communities will form and pursue
the dreams of individualism for many more decades ‑ before they are snuffed
out by an encroaching reversion to primitive tribalism.
[Note: Most of this
chapter was written during the 1990s, before Islamic extremists toppled the
World Trade Centers on September 11, 2001.]
─────────────────────────────────
CHAPTER 13
─────────────────────────────────
THE
TWO CULTURES ‑ PART I
The previous chapter
presents a speculation to account for the rise and fall of civilizations.
I will now consider other factors that may contribute to their fall. In this
and the next chapter I call attention to a troubling situation: there seem
to be two types of people in today's world, and their ways of thinking, and
their values, are the source of "polarizing" conflicts in academia, politics,
foreign relations and everyday life. It may be important to understand the
origin of this two‑sided facet of human "ways of thinking," for, as I claim,
they may play a role in the fall of civilizations.
Review of Brain
Evolution
In this section
I will review material in previous chapters concerning brain specializations,
and present them in a way that serves my present purpose of understanding
why today's civilization appears to consist of a continuum of people at the
ends of which are two distinct and incompatible types. The activities of
people at the extremes tend to produce what has been referred to as "Two Cultures."
If we go back far
enough in our human ancestry, we will find that the left cerebral cortex,
or left brain, LB was identical to our right cerebral cortex, or right brain,
RB. By having essentially identical brain halves our remote ancestors benefited
from a form of redundancy that was valuable in case of injury to one side.
Although a slight specialization for sequential tasks probably developed
in the left brain of our pre‑human ancestors, left brain specializations began
a dramatic evolution sometime in the Pleistocene, possibly 200,000 or 300,000
years ago, perhaps in response to an environmental opportunity presented
by one of the interglacial warmings that have been occurring at approximately
100,000 year intervals for the past half million years. To first order, RB
remained unchanged while LB began to allocate small areas for new, specific
tasks. The ability to talk, as a supplement to gestures, evolved in LB's
frontal lobe (Broca's Area), which was accompanied by the evolution of an
ability to comprehend speech in LB's temporal lobe (Wernicke's Area). Eventually the left side also developed a capability
for logic, a form of sequential thinking that requires the type of neural
architecture involved in language. Logical thought is most effective when
emotional intrusions are minimized. Hence, we may assume that as LB's capacities
for logic evolved it became somewhat disconnected from the limbic system,
where emotions originate.
Modern man's LB
owes its "power" to repeated triumphs of wresting control from RB on matters
that were best performed by newly‑evolving LB modules. Logic is more powerful
than intuition when novel situations are encountered. Novel situations and
opportunities must have been frequent during climate transitions, such
as occurred at the close of the last glaciation, approximately 12,000 years
ago, and during the previous interglacial, occurring 129,000 to 116,000 years
ago.
Whereas brain modules
appear to "work together" to the untrained eye, a more likely dynamic is
that they form an elaborate system of competing modules. Every situation encountered
is "presented" to modules, and any module that "recognizes" the situation
is an invitation for it to become aroused (anthropomorphically speaking);
if the module “recognizes” the situation, it seeks activation from the "reticular
activation system," or RAS. The RAS chooses a "winner" from among the competing
modules (based on past experiences, presumably, as well as inborn predispositions),
and the RAS grants authority to one module. That module is given access
to the frontal lobe modules for formulating behavior and commanding it (via
the motor strip). The winning module’s recommended specific behavior (such
as saying or doing something) is then implemented.
This modular arrangement
makes good sense from the gene's perspective. Indeed, the more one thinks
about it the more impossible any alternative seems to be. Consider any given
brain circuit; there will be one gene that has the greatest effect upon it,
a second gene that has the second‑greatest effect, and so forth. The gene
with the greatest effect on this one circuit is unlikely to affect ALL other
brain circuits with which this circuit interacts, which will also be true
for the other genes that affect the circuit under consideration. Therefore,
if this one circuit works with the others harmoniously it will not be due
to the genes that create the one circuit; rather, it will be due to the
forces of natural selection that pass judgment on genes that create new
circuits (and modify existing ones). The genes for any one circuit do not
"know" about the other circuits, even though they can work harmoniously with
them.
We humans are observers
of the product of the many brain circuits, and we should resist the temptation
to attribute the apparent harmony of mental performance to a harmony of genetic
design. Any harmony that we observe is probably illusory,
since it can be produced, as it most likely is, by a competition of modules
with a continuous unfolding of winners.
Why was LB the site
for mutations conferring the new capabilities of language, logic and other
sequential tasks (if you remember reading this material from a previous chapter,
then skip this paragraph)? One speculation attributes it to a subtle difference
in LB's role for our arboreal ancestors. LB commands the right hand, which
long ago, due to some random selection, took on the task of reaching for
fruit while the left hand stabilized the body by holding onto a branch. Picking
fruit involves a sequence of actions, which led to the development of small
neural regions devoted to each sub‑task and their integration with each other.
Since neural connections within the small neural region relied upon short‑distance
communication, the neuron axons in these regions required less myelinization
(reliance upon a fatty tissue covering of the axon to provide electrical
insulation for better communication to distant neurons). The genes for LB
development became more adept at producing small neural networks that were
less myelinized (having less of the myelin "white matter," thus accounting
for LB's appearance of having a more grayish color). These small neural networks
accomplished specialized tasks with a neural activity that was more‑or‑less
independent of neighboring areas. When an LB neural network area finished
its task it would communicate a "result" to specific other areas, which in
turn also tended to be independent and specialized in their operation. Thus,
LB lost the "holistic" nature of the right, as it specialized in mastering
sequences of specific sub‑tasks that were connected to each other for the
accomplishment of an overall task ‑ such as talking or comprehending speech.
LB's new specializations
rendered it suitable for playing supporting roles when RB recognized specific
situations requiring the specialized performance. LB paid a price for becoming
a specialist. For example, when it assumed responsibility for language it
lost some ability for monitoring body part position in relation to the immediate
physical environment. This task is now performed almost exclusively by RB's
"inferior parietal lobule," a region that is a homologous counterpart to
the Wernicke language area in LB. So, as RB became dependent on LB for language,
LB became dependent upon RB for graceful movement. So
far, in this brief recapitulation of the evolution of LB, we can view the
two brain halves as working together.
RB did not have
to understand how LB did what it was specialized to do (nor could it); RB
merely had to trust it to do what it was good at doing, and RB called upon
it when the situation required. The use of "logic" to understand novel situations
is an LB specialty. RB must be baffled by what the left is doing when it
thinks through a sequence of logical operations. (Similarly, LB would be just
as baffled by how the right side instantly recognizes a face.) Since RB is
a more fully‑connected neural network system, it would seem to be well suited
to recognizing when a situation requires LB help. When RB makes this determination,
it hands control to an LB region for the duration of the specific task. At
least this may be the way things started out when the LB specializations first
began to evolve.
When a human invents
a new tool, he uses it for pre‑existing purposes. However, the tool is also
available for use to accomplish other tasks, perhaps tasks that existed before
the tool existed. I view the new LB specializations in the same way. The
brain did not evolve to comprehend reality; rather, the little reality that
it does comprehend, between the many distorted comprehensions, is due to
the fact that a better understanding of some realities were useful to the
survival of our ancestor's genes. We can speculate that the human brain's
greater ability to understand the world around it is an unintended consequence
of an original need to perform specific sequential tasks. For example, the
rewards of allocating the right hand for the task of reaching for fruit led
to the evolution of sequential structures in LB, which allowed language to
develop, which led to the making of sophisticated tools, which helped to create
categories for placing and using words, which is the basis for abstract thought,
which may eventually allow humans to achieve liberation from their genetic
enslavement. What a wonderful outcome of the prosaic task of reaching for
fruit.
My assertion that
the sequential abilities of LB were used by RB to accomplish long‑standing
tasks more successfully is consistent with the notion that the cerebral cortex
itself, both left and right sides, is used by sub‑cortical structures for
the more effective accomplishment of pre‑existing goals of survival and
reproduction. Thus, the limbic system mobilizes the more recently evolved
cerebral cortex by “driving it,” invoking emotions as necessary, to engage
the world in ways that lead to the creation and raising of offspring carrying
the genes that assembled the individual and his close relatives.
The brain was not
intended to aid in comprehending reality, per se, and the
genes can be excused for not anticipating that LB would someday figure out
that the individual is enslaved by its genes – a central message of this
book. This unintended achievement is due to LB's prefrontal cortex, the part
of the frontal lobes forward of the motor strip. Both the left and right
prefrontal cortices have grown in size at an amazing rate in relation to
the rest of the brain, during the past half million years. Interestingly,
the prefrontal areas do not contribute to the component of intelligence measured
by IQ tests. Rather, the prefrontal cortex works with the limbic system to
produce elaborate planning capability, or "executive function." It is likely
that “what if” scenarios are orchestrated by the prefrontal cortex, and are
evaluated by the underlying limbic system ‑ to assure that genetic goals are
being served when a plan is adopted. I speculate that LB is less restrained
by the limbic system, in order to rely upon logic when creating "what if"
scenarios, and to subject these hypothetical scenarios to objective criteria
for establishing the truth of a novel situation.
Because LB is better
suited to comprehend situations requiring logical analysis, it would have
frequently found itself in situations where it recognized a problem that
RB was totally unaware of. LB needed to take action, yet RB was unable to
endorse LB's call for action since it did not "comprehend" the problem. To
the extent that survival (of the genes) depended upon LB being allowed to
act when RB didn't comprehend the need for it, there would be a need for some
level of trust in LB's compelling request to take charge in dealing with
a matter that only it could handle. The trust would have to reside in both
RB and the reticular activating system (RAS). Perhaps it became more efficient
for LB to initiate actions without RB's consent, with a direct appeal to
RAS. This would eventually lead to trouble for RB.
In a primitive setting
there may have been very few situations requiring LB's specialized understanding.
However, the more capable that LB became, the more the new world created
by LB needed LB solutions. LB’s new world included sophisticated weapons
and tools, horticulture, animal domestication, and the formulation of logical
strategies for both everyday living and inter‑tribal warfare. RB would be
calling upon LB to act more and more, and allowing LB to unilaterally decide
to act for a growing number of situations in the emerging new world of LB's
creation. It is inevitable that as LB assumed more initiative in recognizing
situations that needed LB attention, LB would have assumed responsibility
for responding without consulting RB. The harmony that existed between RB
and LB was at risk of being upset, the more so as the new "upstart" LB assumed
more roles and a greater importance in surviving the challenges of the late
Pleistocene. Occasional LB/RB disputes of authority may have become common
during the Holocene.
The disputes would
not be between the entire LB and the entire RB; they would be between specific
parts of LB and a large, diffuse part of RB. Recall that RB is a more fully-connected
neural network, whereas LB consists of many specialized, task‑specific neural
networks. RB recognizes situations from the totality of the sensory input,
but isolated modules within LB recognize situations needing LB attention.
Since niches in
the tribal setting tended to be of either an artisan type or the non‑artisan
type, I believe that eventually people were "born with a predisposition"
to occupy one of these two niche categories. It is quite possible that most
people are born with the ability to take a reading of the environment and
develop in the direction of one or the other niche type. It would make sense
if the brain circuits that "take readings" of the environment, and hence of
the relative merits for differing ontological development paths, also take
into account the strengths (predispositions) that the individual inherited.
(This whole process in which an individual's development unfolds in some way
that approximately optimizes its usefulness to the genes within is a daunting
subject, but it must be studied by future generations of sociobiologists.)
In this way, I suggest, individuals are born with a predisposition for developing
into one of two types: the sequential‑style artisan type, or the holistic‑style
primal type. The rest of this chapter is concerned with the differences
between these two types of adults and their implications for the fate of
civilizations.
RBS and LBS Defined
In English there
has been a longstanding use of the terms "head" and "heart" to signify two
distinct ways of thinking and of "being." There is an unending flow of movies
and books based on the conflict of the head and the heart, so since the terms
are in such widespread use it should be unnecessary to define them. However,
it is now obvious that the common term "head" is somehow associated with
the perspective of the left brain, LB, while "heart" is somehow associated
with the perspective of the right brain, RB.
I am tempted to
present specific speculations about the neural mechanism that accounts for
this dual association. For example, I could suggest that the "head" person's
LB dominates his RB, forcing RB to play a supporting role; and that the "heart"
person has an opposite dominance/support role relationship. Or I could suggest
that the "head" person's LB asserts itself without RB permission, like an
intrusive, uninvited guest, whereas the "heart" person's LB is not assertive
in this manner. As an alternative, I could allege that the "head" person's
RB is more similar to an LB, and the two hemispheres work together because
they do not have conflicting values and life agendas; whereas the "heart"
person has two RBs that work together in similar harmony. As a final speculation,
which I tend to favor, I could invoke the prefrontal cortical areas, and
state that the "head" person's LB prefrontal cortex has a stronger inhibition
of the RB prefrontal cortex compared to the opposite inhibition, rendering
LB's more closely connected LB posterior lobes a greater "voice" in perceiving
the world and determining behavior; whereas the "heart" person has an opposite
prefrontal lobe inhibition pattern, giving the RB posterior lobe perceptions
of the world greater voice and a stronger influence over behavior.
I will not take
a position on these alternative theories, as I believe the neurosciences
are not yet advanced enough to warrant a preference for any of the foregoing.
Indeed, perhaps none of the hypotheses I've identified are true. Or maybe
several apply. It is not necessary for us to know why so many people tend
to fall into the "head" and "heart" categories in order for me to proceed
toward an important goal of this book. I merely ask the reader to accept
that with some people it is possible to categorize them as tending to belong
at the "head" end of a spectrum of traits, while others belong at the "heart"
end of the same spectrum, acknowledging that most people are somewhere between
the two extremes. (Anyone reading this far is a “head” person; the heart
people would have discontinued in disgust during Chapter 1.)
I also ask the reader
to consider the possibility that some people are born with a predisposition
toward one or the other type, or with a predisposition to be a specific blend
of the two extremes. And finally, I ask the reader to accept that those
at the "head" end of the spectrum are a product of recent evolution, meaning
that people having the "head" predisposition were less common the farther
back in our ancestry we go. This last assertion is essentially a "thought
experiment," since I cannot think of any way to prove it using data from
the archeological record. The last point also has a corollary, worth stating,
which is that "heart" people have "always" existed. Again, the archeological
record probably cannot be used to support that assertion. However, I think
that both of the alleged head/heart trends over evolutionary time are reasonable,
possibly self‑evident, and I shall risk making no further argument to support
it as I proceed with my larger argument.
I am going to use
the terms RBS and LBS as shorthand for "Right Brain Style" and "Left Brain
Style" to refer to the beliefs and ways of experiencing life by the "heart"
and "head" people. By my subjective reckoning, LBS people are much rarer
than RBS people, and most people are a mixture of the two extreme types.
To estimate their numbers it will be convenient to make use of readily available
polls of religious belief. I shall use "disbelief in God" as a proxy parameter
for identifying LBS people (which admittedly neglects culture's influence
upon an individual's religious beliefs). One could also use "disbelief in
immortality, or spirits, or astrology" but these questions are less frequently
included in polls. Based on the frequency of atheism in the American population,
only 4% of people are LBS (and 0.8% are hard‑core
LBS). The 4% estimate may strike some as too small; I agree, and cite that
since the atheism rate in Europe is greater than
in the United States the 4% value has
probably been influenced by the traditional strength of religion in the
U.S. from the country's
founding (Hofstadter, 1969). Perhaps 5 to 10% would be a better estimate
for the LBS rate among the Western world's population. Among elite groups
of professionals the LBS rate is higher. For example, among physical scientists
who are members of the National Academy of the Sciences,
the rate of disbelief in God rises from the general population's 4% to an
impressive 79%! (Larson and Witham, 1998).
In order to describe
a conjecture on the evolution of the human LBS/RBS distribution I will describe
how one might measure an individual for the purpose of scoring him for placement
on an LBS/RBS spectrum. Let us begin with IQ tests to illustrate how a person's
LBS/RBS score might be measured. Every IQ test that I'm aware of has many
subtests, and these are usually averaged in two groups. The two groups were
devised before LB/RB studies showed that brain competencies are lateralized.
Nevertheless, it is clear that one part probes mostly RB posterior lobe
function while the other probes mostly LB posterior lobe function. For example,
the "verbal" and "performance" scores for the WAIS and Wookcock‑Johnson tests
are crude measures of competence of the posterior lobes of LB and RB, respectively.
With sufficient motivation I am sure that someone experienced in psychological
testing could devise an objective measure that scores how LB and RB posterior
lobe competencies are employed by the frontal lobes to produce the subjectively
recognized RBS and LBS individual. In addition to a reworking of existing
subtests, such as the WAIS and Luria Neuropsychological Investigation, I
would suggest including questions about belief in angels, ghosts, spirits,
communicating with the dead, telepathy, precognition, prayer, voodoo, astrology,
touch therapy, life after death, and God. Assuming an effective LBS/RBS test
could be devised, it would then be possible to imagine the "thought experiment"
of testing a population of people from different times in our evolutionary
past.
Evolution of LBS
If pressed to estimate
when LBS people began to appear, I would say approximately 200,000 years
ago, which is before the split of the three principal races of man: African, Asian and Caucasian (Rushton, 1995). Before then,
everyone was all "heart." It may be worth illustrating how the population
distribution across an LBS/RBS spectrum evolved from before the appearance
of brain lateralization to the present. I rather think the change was not
a "shift" but rather the rise of one "wing" of a distribution. The population
distribution change would have been "driven" by the existence of niches that
were newly within reach of being occupied by an otherwise slowly evolving
human species.
The following two
figures are meant to illustrate an imaginary experiment in which our primitive
ancestors are measured by a LBS/RBS test and scored on a LBS/RBS spectrum.
Figure 13.01 shows what might have been measured prior to the exploitation
of artisan niches, let us say 300,000 years ago,
before our cerebral hemispheres were greatly lateralized. At that time the
only niches being exploited required "LBS minus RBS scores" in the 70 to
90 point region, which in this illustration is well matched by the population
distribution.
Figure 13.02 shows
a hypothetical population distribution after new niches appeared and began
to be exploited by a subset of the population. The new niches might have
been the use of language in hunting, toolmaking, horticulture, animal domestication,
animal breeding, construction of food cache buildings, or anything requiring
nearly full‑time dedication and requiring conceptual thought or sequential
thinking modes. I have indicated that the population distribution is not
symmetrical, but has a bulge at the high "LBS minus RBS" end of the spectrum.
When comparing individuals from the two ends of the spectrum there would be
a stark contrast, and it would be tempting to say that they represent two
distinct "types." The population distribution does not have to be noticeably
bimodal for this impression to exist.
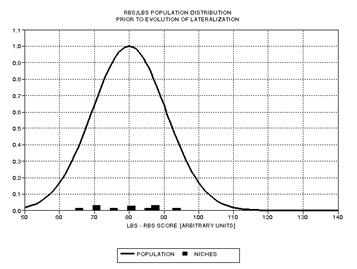
Figure 13.01 Hypothetical
distribution of primitive population on an imaginary LBS/RBS spectrum. The
boxes along the baseline represent niches that exist and can be feasibly
exploited at this stage of human evolution.
Figure 13.02 Hypothetical distribution after new niches
appear, perhaps associated with a climate change, showing how the population
distribution might change in response to the new opportunities for niche
exploitation. The right-most symbols represent "artisan" niches requiring
a sequential type of thinking that only a left brain can perform.
I suspect that if
a valid LBS/RBS test could be devised, results from today would produce a
distribution resembling this figure. The two cultures would be represented
by scientists, engineers and lawyers having jobs located by the right-most
symbols, whereas salesmen, entertainers and the rest of the population would
have jobs represented by the other symbols.
The conditions that
favor the evolution of LBS are cultures rich with niches for the analytically
inclined, which are abundant in today's technical world. In previous generations
a Bill Gates would have been a lackey to a king, an artisan “on call” for
odd jobs the king assigned. Just as people are born with different abilities
for scoring well on IQ tests, it is inevitable that people are born with
different predispositions for being LBS versus RBS. I believe that some people
are thus pre‑adapted to become LBS, and prosper within a culture that rewards
LBS, whereas in others the vast majority would be destined for life as RBS,
or something in between.
Those people who
are genetically predisposed to being strongly RBS will be forever resentful
of the minority of strongly LBS people among them, especially if the culture
rewards LBS talent as much as it does today. The vast majority of people
will merely feel "uneasy" about the presence of strongly LBS people. This
inherent resentment of one type for the other is hinted at in C. P. Snow's
book The Two Cultures (1961). It is also possible to gain
insight into the dilemma posed by LBS individuals in a predominantly RBS society
by reading between the lines in Resentment Against Achievement (Sheaffer, 1988), Anti-Intellectualism in America (Hofstadter, 1969)
and The Great Roob Revolution (Price, 1970).
In today's Western
culture, dominated by the products of LBS achievements, it is easy to portray
primitive people as primitive. Such pronouncements may not be welcomed by
the RBS majority, for they recognize a resemblance of their outlook with
that of the primitive. For example, RBS people tend to see all things in terms
of the animate, whereas LBS people tend to see things in inanimate terms
(cf. Ch. 2). The primitive, who is by inclination an adherent of an RBS world
view, imagines that spirits reside in the trees, the wind, lightning, and
all other natural phenomena. They conceive of the "forest as parent" and
"a giving environment, in the same way as one's kin are giving" (Bird, 1990).
The Inuit "typically view their world as imbued with human qualities of will
and purpose" (Riddington, 1982). American Indians in pre-Columbian times
also viewed the world as controlled by spirits (Aleshire, 2001).
In contrast, the
scientist has cleansed his world view of these ancient spirits, and he tries
to perceive things as mechanistic (this attribution is for the hard‑core
LBS scientist, who is more likely to be found among the elite, high‑achieving
scientist population). The hard‑core LBS person is a reductionist, who automatically
perceives all phenomena as the inevitable unfolding of a mechanistic universe
(only a minority of high‑achieving scientists would object to this characterization).
If you like what you're reading, then you, like the author, are extremely
LBS.
As an aside I will
present an amusing example of the differences between the approaches to understanding
something by an LBS and an RBS person. It occurred at a conference on "Imagination
and the Adapted Mind: The Prehistory and Future of Poetry, Fiction and Related
Arts," UCSB, August 26‑29, 1999. The invited speakers seem to have been
intentionally drawn from the core of the opposing “two cultures” camps.
It was amusing for me to watch a hard‑core RBS participant's consternation
over the fact that inanimate objects enter our mouth, and inanimate objects
leave at the other end, yet "we ourselves are animate." For him that was
a dilemma, requiring appeal to the magic of words, which he provided, referring
to "transubstantiation" ‑ as if this magical invocation explained something
and solved his dilemma. But it is only a dilemma for the RBS person. For
me, a hard‑core LBS person, all so‑called animate objects are more accurately
to be thought of as "inanimate automatons." We are composed of the same atoms
as everything else, and no magic transubstantiation occurs when the atoms
of food and water enter or are incorporated into our bodies. Those atoms
are subject to the same laws of nature that moved them before they became
part of this exalted entity we call human. The RBS person, being unaccustomed
to thinking of the human body as a mechanism, would naturally be troubled
by this trivial phenomenon.
The modern world
rewards left‑brained styles of thinking more than at any previous time. The
LBS people of today appear to be "pre‑adapted" to our times. But pre‑adaptations
do not exist by intention. Either they are accidents, in which a small
opportunity is exploited by a novel mutation, or a wide niche is exploited
gradually by a series of mutations, each adding an increment of capability
to the first. I am wondering if the gradual evolution of LB specializations,
which underlies lateralization, is in fact a case of the latter. For example,
imagine an evolutionary scenario in which a sequential brain circuit enables
a toolmaker to produce better spearheads than his brothers, and his genes
are rewarded. Next, that line of men who have the slightly more sequential
brain circuit in their LB, which the tribe may designate as their official
toolmaker, undergoes another mutation that further improves his toolmaking
ability ‑ he might simply demystify the stone that he works with. By this
process he may lose the belief that the stone has feelings, and is hurt when
struck, or that the stone wishes to be elsewhere, or that the stone will
invoke the spirits of his stone relatives to wreak revenge upon the heartless
chipper ‑ and this is an asset for performing his toolmaking task. We now
have someone who not only tends to depersonalize the stone he must work with,
which frees him to handle the inanimate stone better, but a person who tends
to demystify in general. He is more likely to perceive the wind as really
not caring about humans. He might be inclined to discard the many spirits
that so preoccupy his fellow tribesmen - and if he is smart he will keep
these new beliefs to himself.
What I am proposing
is an evolutionary selection and maintenance of a specific type of variation
within a species, for there are reasons the majority of tribesmen should
maintain their spirit‑styled outlook. Spirits, I shall assume, are a short‑hand
way of enabling people to deal with each other, and the majority of tribesmen
are faced with having to compete with their fellow men rather than the stones
to which the lonely toolmaker is relegated. I view the toolmaker as heralding
an era of the division of labor. He is the prototype for the artisan, who
occupies the broader tribal niche of improving weapons, inventing new ones
(like the bow and arrow), refining horticulture, building food storage huts,
domesticating animals, inventing the pastoral way of life, and later, irrigation,
wheeled carts, atomic bombs, the internet and nanotechnology. The opening
up of artisan niches, starting with the toolmaker, provided ever greater
rewards to the man who viewed nature as spiritless, or inanimate. The human
answer to this ever‑growing opportunity was to invest more in left‑brained
circuitry specialization for dealing with the inanimate world. These lucky
new people may have paid a price in being less apt in dealing with human relationships,
but this would often be mitigated by a tribal understanding that these people
were "different" and should be left alone to do their artisan work. They
may have been looked upon as childish, or unmanly, since they were probably
exempt from pillaging and other proud warrior exploits. The tribal chief
might have thought of the artisans the way people later thought of the herds
of animals that they domesticated: they had to be "kept" because they were
useful to the tribe.
This speculation,
that some people are naturally LBS, is meant to explain why the two cultures
phenomenon is so pronounced. It suggests that some people are fated at birth
to occupy either one of the small but growing number of LBS niches, or instead
to occupy the many though decreasing number of RBS niches. Other people,
being only partly inclined to artisan niches, could go either way in their
personal development, and they would be wise to take a reading of the likely
rewards for competing career paths when they make their (unconscious) life's
path decision.
The Threat of Individual
Liberation
The evolution of
capable prefrontal lobes, and a newly‑fashioned LB specialized for analytical
insight, obviously helped our ancestral genes survive. This much can be safely
concluded by merely citing that we have the hardware in question. But new
tools can sometimes cut in two directions, metaphorically. A brain that
can put two and two together can theoretically put itself to uses that are
subversive from the perspective of the genes; such an individual is at risk
of liberating himself from the enslavement of "being a tool for the community";
he may think taboo thoughts of "becoming the captain of one's own ship, enslaved
to no one." The toolmaker is at risk of “walking away”
from his assigned role as a tool for the tribe!
A creative prefrontal
lobe might ponder scenarios in which the individual essentially "thumbed
its nose" at the tribe by going off into the woods to live a simpler life?
Before the creation of super-tribes, which offered specialized niches for
strangers to fill, this way of thinking would have been suicidal. But during
the past 12,000 years, during our present Holocene interglacial, some options
for walking away from the genetic agenda have become feasible, and should
be a real "concern" to the genes.
Let's review the
main theme of this book, as presented in the Introduction. For each hypothetical
behavior that has an innate component, we are to make an evaluation from
the perspective of the genes, and then make an evaluation from the perspective
of the individual. We must invent the concept "individual welfare," or "individual
best interests," to continue this experiment. I shall infer its essence
by presenting just two brief examples of thought experiments to illustrate
the concept "outlaw genes." A larger set of examples, with fuller explanations,
is presented in Chapter 13.
The individual who
invests in raising offspring is embarking upon a long‑term and exhausting
life course. Sex leads to babies, and for women this means the commitment
of a life of parental enslavement. For "dutiful" fathers, sex and babies
can lead to a similar enslavement. If a man or a woman gave thought to this
situation, and instead decided to pursue an individual‑fulfilling life of
aesthetic pleasures ("smelling the roses," watching sunsets, appreciating
music) that person could in theory lead a less exhausting and more tranquil
and fulfilling life.
For my second example,
consider the young man who is expected to participate as a warrior in raids
of neighboring tribes (or nations, if our locus is the Holocene). Marauding,
pillaging and engaging in battle are dangerous activities. The individual
who says "no" to tribal offensive campaigns, thereby flaunting unthinking
patriotism, and who can successfully avoid combat without paying the price
of humiliation, would be safeguarding his best interests.
Let us now pretend
that we can calculate outcomes of various actions, and ascribe to them some
quantified measure of Individual Welfare Value, IWV. Let us also suppose
that we are able to identify how strongly each gene contributes, under average
conditions, to eliciting each action. I have in mind a model for generating
behavior in which a person's genotype interacts with his Environment to produce
an expressed Phenotype, or GEP, a process best described by Symons (1979)
and treated in Chapter 6. By adopting some average environment we can in
theory calculate 1) the probability that a given gene will elicit the action
under question, through the GEP analysis, and 2) the effect of this action
upon the frequency of genes in the future genome. This allows us to link
an action to a measure I will call Genetic Survival Value, or GSV. If we
can do these things, then we can plot a point corresponding to that gene
in IWV and GSV space. To wit, refer to Fig. 12.03, first presented in the
Introduction and reproduced here.
Since the concept
of "outlaw gene" is so important, its explanation is worth repeating. In
the figure we have four quadrants. The upper‑right quadrant is where the actions
elicited by most genes reside, as they provide value to both genetic survival
and individual welfare – such as breathing and eating. The lower‑left quadrant
contains mutant mistakes, and genes that elicit such actions should survive
no longer than the individuals who carry them. Genes in the upper‑left quadrant
are unlikely to exist, except by mutational accident, since there is no way
for Nature to code for an action that destroys the code. The most interesting
quadrant is the one in the lower‑right. The actions found here are actions
that should be selected by Nature just as strongly as for those in the upper‑right
quadrant since they benefit the genes that code for them; but the actions
in this lower‑right quadrant exist in the face of harm done to individual
welfare. This is the quadrant that motivated my interest in sociobiology
on February 23, 1963, when I first created
the figure.
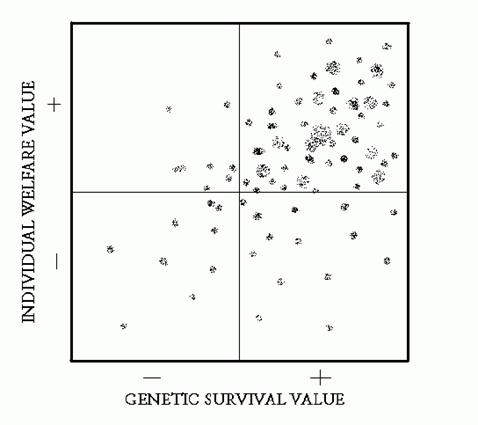
Figure 13.03. Gene Value and
Individual Welfare scatter diagram.
The "outlaw genes"
are the ones found in the lower‑right quadrant. There is no way for natural
processes to eliminate outlaw genes. Indeed, natural processes will reward
them when they mutate into existence, and if they conflict with other genes
a balance will naturally be struck. The balance could involve some form of
accommodation, such as making the individual believe that the actions in
question are attractive. The more common solution, however, is for the genes
to invent emotional rewards that are meant to overcome any logical hesitation
that might exist by individuals who are otherwise unconvinced by the instinctive
imperative. Short of taking evolution into our own hands, humans will be
burdened forever by the "victimization" that outlaw genes perpetrate upon
the individuals they make.
An LB individual
might chance upon the thought that some things everybody does are not in
a person’s best personal interest, and if his social setting provides a way
to avoid the unwanted action he might actually plunge into self‑liberation
by resisting the instinctive, self-defeating act. For example, a woman might
choose to not have children, or a man might find a way to avoid going to
war. Horrors, if the genes allowed these acts of individual liberation to
continue! So, given that liberated people would have fewer offspring, we should
expect to see some evidence that a tendency to think liberated thoughts would
be discouraged by the genes, or that thinking the way the genes "want" us
to think would be attractive. Is there any such evidence?
The Two Cultures
Anyone who today
reads C. P. Snow's The Two Cultures and the Scientific Revolution
(1961) would be struck by resemblances of Snow’s descriptions to LB and RB
styles of thinking. During the 1980s neuropsychologists were making headlines
for research that revealed LB/RB differences, due mostly to the study of
split‑brain patients who had their interconnecting corpus callosum surgically
cut to control epileptic seizures. Snow contrasted "literature academics"
with scientists, as they seemed to epitomize the extremes of a continuum based
on his personal experience. His work during previous decades had placed him
in contact both groups, and he became fascinated
by their differences and their inability to communicate. Not only could they
not communicate, they didn't want to. It was mostly the literati
who shunned the scientists, and made disparaging remarks about them, while
the scientists went about their work full of optimism that their importance
was on the ascendance and they were eclipsing their rivals.
The word “rivals”
is apt, because, as the previous sections of this chapter have made clear,
I claim we are really talking about people who think with an LB style versus
those who think with an RB style, which are products of rival brain circuits.
The community of RBS intellectuals have frequently "made fun of" LBS people,
as if they needed to discredit them in order to compete with them. I recall
from my college days a prankster's sign posted on an arch of an engineering
building that read "All my life I wanted to be an engineer; and now I are
one!" At the time, I realized the point, and found it funny in a bittersweet
way (I was enrolled in the School of Engineering at the time). But
now I have a better appreciation of the motives for the humor.
The field of anthropology
was "hijacked" by Franz Boas early in the 20th Century. His student
disciples, Margaret Meade, Ruth Benedict, and others, worked with Marxist
zeal to discredit the influence of genes on behavior. The main determinants
of behavior were supposed to be the wretched influences of a corrupt society acting upon an innocent and malleable
human nature. Anthropology is still undergoing turmoil, as the forces of
rational LBS people seek to rescue it from a century of RBS abuse. The apologists
for the RBS suppression of nature‑oriented investigators have occasionally
revealed their motivation; they were afraid of the political ramifications
if it was generally believed that badness in people is inevitable.
The latest RBS campaign
to discredit LBS thought has various names, the most common being "political
correctness," "deconstructionism" and "postmodernism." Again, there is a
Marxist heritage to these new thought enforcers (Ellis, 1997; MacDonald,
1998). Their argument hinges upon a grain of truth, but exploited beyond
reasonableness. At the most fundamental level, so the argument goes, all
things are inherently meaningless and without value, and anyone who espouses
a belief is deluding himself that his belief can be measured against someone
else's belief. Hence, science is just one more pompous and fashionable belief
system, and it is used to hoodwink the general public. Consequently, someone
from the humanities should feel free to challenge anything sacred in science,
such as gravity, the conservation of energy, and all manner of physical law.
And therefore all LBS people are charlatans! Yet, amazingly, their RBS beliefs
somehow escape their "fundamental meaningless law," and RBS beliefs remain
unscathed. Therefore, all RBS people are Truth Tellers! Ellis (1997) states
that "In the span of less than a generation, university humanities departments
have experienced an almost unbelievable reversal of attitudes, now attacking
and undermining what had previously been considered the best and most worthy
in the western tradition." It is fitting that the most corrupt field of
study is "literary criticism," the field that C. P. Snow chose as exemplifying
the culture opposed to science. The demise of the humanities during the second
half of the 20th Century is so bizarre, and out of control, that it's an
easy subject to get worked up over. I shall refrain from the temptation, in the interest of pursuing a bigger subject
(see Weinberg, 2001 for science-based rebuttals to postmodernism.)
It is no coincidence
that the early 20th Century hijacking of anthropology, and the
late 20th Century hijacking of humanities departments, has been
inspired by Marxist intellectuals. Marxism and communism are political expressions
of "collectivism." Individualism has been in conflict with collectivism for
as long as human history has been recorded. The Eastern cultures are strongly
collectivist. It should be no surprise that China embraced communism,
and remains today the staunchest holdout and defender of communism. Remember
that the LBS individual is prone to think subversive thoughts, and discover
new ways of viewing the underlying motives for the collective force of societies.
I claim that the genes are most likely to find the RBS‑prone person a fitting
candidate for enforcing the collective (genetic) agenda by monitoring the
errant wanderings of LBS intellectuals. Sensing that RBS vigilantes are watching
them, LBS interlopers are always looking over their shoulder for the enforcers
of gene interests.
High achievers have
probably always sensed a resentment of their successes by those less able
or less motivated. Robert Sheaffer (1988) wrote Resentment Against Achievement: Understanding the Assault Upon
Ability to document this interesting social phenomenon. He writes that
"Throughout recorded human history the ebb and flow of the love of achievement
and the resentment against its successes have been major forces behind the
rise and fall of civilizations and empires… Ahievement‑oriented values like
tolerance, liberty, and the freedom of the individual to work hard and
enjoy the fruits of his labor provide the motivation necessary for a civilization
to grow and flourish."
A superficial assessment
of how the genes should perceive achievers might challenge my thesis that
the resentful are working on behalf of the genes. But consider that what
the achievers are really resenting, and the fear the genes are really responding
to, is that the achievers are those unpredictable LBS artisans who can't
be trusted to stay loyal to the tribe. A person who leaves the service of
the genes to pursue individual goals is not an asset to the genes; rather,
such a person is a threat to the genes because his example might inspire others
to do the same, and thereby threaten a wholesale abandonment of the collective
genetic agenda. Lo to him who dares write a book urging individuals to seek
liberation from their genetic enslavement!
─────────────────────────────────
CHAPTER 14
─────────────────────────────────
TWO
CULTURES ‑ PART II
"The whole thing
is so patently infantile, so incongruous with reality, that to one whose
attitude to humanity is friendly it is painful to think that the great majority
of mortals will never be able to rise above this view of life."
Freud, Civilization and its Discontents (1930), commenting on religion.
Examples from Everyday
Life
Sometimes a theoretical
argument is best illustrated by examples, so I want to start this chapter
with the following three newspaper articles.
Item 1. On October 31, 1999, Egypt Air Flight
990 crashed shortly after taking off on a flight from New York to Cairo. The cockpit voice
recorder and flight data recorder indicated that a lone relief co‑pilot
was at the controls at the time the tragic event began to unfold. The co‑pilot
said the Arabic equivalent of "I put my faith in God," then disconnected
the autopilot, pushed the nose down, and sped toward the ocean below at Mach
0.9. The pilot returned to the cabin, asked "What's going on?" He took his
seat, and pulled up on his control yoke while shouting to the co‑pilot to
"pull with me." But the co‑pilot persisted in pushing down, which dis‑engaged
the tail elevator's coordination mechanism, causing the elevators to go in
opposite directions, which made the plane start to spin. During the dive
the engines were turned off. It crashed, killing all 217 aboard. This information
was recovered and reported by the American FAA about 3 weeks into their investigation,
and implied that the co‑pilot may have committed a mass murder suicide.
However, in Egypt the public reaction
was one of disbelief. The Egyptians suggested that the Americans had engaged
in a cover‑up to defend the reputation of an American airplane manufacturer
(Boeing) and a desire of the Americans to discredit Egyptians. American
newspapers began reporting on "the cultural chasm between the two societies."
The Egyptian newspaper Al Akbar described the American interpretation thusly:
"This hallucination might be accepted in an American movie. But it is difficult
to be convinced of this [being done by] a mature and sensible Egyptian."
An Egyptian magazine editor summed up the different philosophies: "The American
has learned to conquer life and put trust in science and technology, while
the Egyptian has learned [that] time is more powerful than the human being,
and we cannot stand alone, but that it is better to have God by your side."
My summation would be that "The Americans have learned how to conquer life
by relying upon the left brain to use science and technology to understand
and live effectively in the world, while the Egyptians have continued to
view the world with the right brain perspective in which God is one’s guide
for serving the genes."
Item 2. The Santa Barbara News‑Press reported (1999.02.07) that "LightShift
2000" wants everyone to "meditate for planetary harmony" at noon on the first day
of every month. They claim that synchronized meditation is “a catalyst for
positive energy” that will affect the surroundings. They believe that "silent,
compassionate meditation can shift the collective consciousness of the world,
reverse our course of destruction and truly create positive healing change
in the approaching millennium. ... If you get people banding together and
create a chain reaction of positive energy, that
... creates the light shift." The movement was instigated by a book written
by astrologer Ken Kalb, residing in Summerland, CA (near Santa Barbara) titled LightShift 2000 ‑ Let's Turn on the Light of the World, which
has sold 12,000 copies so far. A LightShift web site (www.lightshift.com)
has been visited 1.3 million times. The stated goal is for a 6 million person
mass meditation January 1, 2000, at 12:12 AM. Somehow they
determined that if only the square root of any population would synchronously
meditate, the overall quality of life would improve. According to my math
this means they need only 78,000 people to save 6 billion, not their 6 million.
I guess math isn't their strong suit. Even if you're "at work or in a meeting,
a short prayer is better than nothing. Or just repeat the affirmation: 'May
peace prevail on Earth.'"
Item 3. Some people are
an embarrassment to the human race! They keep coming up with weird beliefs
that defy logic, which causes the rest of us to conclude that they are “logic
challenged.” One example of a recurring belief is that God will put gold
crowns on the teeth of the faithful "as a sign." After attending a revival
at a church where the magical appearances are reported to have occurred the
faithful will check their mouth, and behold, gold crowns, plates bridges and bands materialize. This phenomenon
first appeared in Argentina in the 1980s, and
spread to Mexico, South Africa, Canada, Britain and now America. The claims fade
after several years, then reappear. A Canadian
went to his dentist to prove his claim, and the dentist reported that he
did the gold work 10 years earlier. When this was reported in the newspapers,
the gold teeth reports mysteriously stopped. "Faith springs eternal" because
it is in our genes. Fortunately, not all humans have it in the same amount,
and those who lack it are blessed with a modern left cerebral hemisphere.
Empowerment of the
Masses
The 20th
Century LBS (Left‑Brain Styled) scientist is acutely aware of the threat
posed by the newly‑emboldened common man, who is preponderantly RBS and considers
his opinion on scientific matters to be just as valid as the expert's. Keay
Davidson (1999) captured this concern using the example of Harlow Shapely,
a Harvard astronomer who opposed pseudo science. Shapely was especially irked
by the readiness of the Macmillan publishing house to publish the book World's in Collision by Velikovsky (an RBS exemplar). Shapley's
position was summarized by James Gilbert (1997) in the following manner: Science "...by its very nature should never be molested
by popular belief; it is the sole purview of those who understand it."
Keay contrasts this position with that of William Jennings Bryan, who believed
in "the democratic community's right to decide the validity of scientific
theory" (Gilbert, 1997).
Jose Ortega y Gasset
wrote in his 1930 book The Revolt of the Masses that the
unschooled common man began, in the 19th Century, to view his
opinion on all matters as having equal validity to those of scholars who
devoted their life to the same matters. Part of this new boldness may be
due to economic growth that brought increasing wealth to the masses; the
new boldness was also nurtured by the entitlements that a democracy creates.
But the mass man's mistaken belief was outwardly attributed to the conviction
that Truth could be found by looking inward, with earnestness and faith.
This subject is treated in greater length in the next section.
Oriental Rejection
of LB
Oriental philosophies,
such as the Tao, eschew LB‑styles of thinking, and attempt to discredit it
in their philosophies for thinking and being. In their place they counsel
ways of thinking and being that are transparently RB‑styled. I will assume
that the reader is familiar with the late 20th Century neuropsychological
findings on brain laterality, as reviewed in Chapter 7. During the 1980s
there was so much reportage in the popular press about brain laterality findings,
many of which were indeed astounding, that some purists referred to the phenomenon
of an over enthusiastic embrace of these new ideas by coining the term "dichotomania."
During the 1990s the purists effectively squelched those inclined to even
a reasonable version of dichotomania, so the exploration of RB versus LB
issues was "still‑born." Be warned; I am a dichotomaniac!
The Tao complains
that "we have learned to put excessive reliance upon central vision, upon
the sharp spotlight of the eyes and mind..." and that "we cannot regain the
powers of peripheral vision unless the sharp and staring kind of sight is
first relaxed." Alan W. Watts (1957, p.19). A
neuropsychologist would immediately recognize the brain laterality connection
of this thought. The old RB is continually monitoring sensory input for signs
of danger, and it does this in a computationally fast and subconscious method,
relying upon parallel processing, comprised of interconnected neural networks.
When the conscious mind focuses upon something, it most‑often does so under
the direction of LB, and uses the high resolution central visual field for
this task. It is uncanny how the Oriental has captured this signal difference
in left/right roles for looking at the world, and prefers one over the other.
The I Ching encouraged
the Chinese mind to arrive at "decisions spontaneously, decisions which are
effective to the degree that one knows how to let one's mind alone, trusting
it to work by itself." (Alan W. Watts, op. cit., p.19). Again, this advice
essentially counsels a person to "turn off" LB, and trust RB to make decisions.
(This is remindful of the advice to tennis players,
golfers, businessmen, etc. presented in a series of The Inner
Game of... books by W. Timothy Gallwey, starting in 1974).
Alan Watts describes
Chuang‑tzu's advice "The perfect man ... grasps nothing..." Also, "it 'fuzzes'
itself a little, to compensate for too harsh a clarity." Later, Watts writes "...both
Tao and Confucius thought that the natural man is to be trusted." (Note
that "natural" must refer to primitive, RBS man.)
In Oriental thinking
there is a strong resentment of LB intrusions. They extol the virtues of
a form of unconsciousness, something "exponents of Zen later signified by
wu‑hsin, literally 'no‑mind,' which is to say, un‑self‑consciousness. It is
a state of wholeness in which the mind functions freely and easily, without
the sensation of a second mind or ego standing over it with a club." (Alan
W. Watts, op. cit., p.23). The term "second mind" that Watts uses refers, quite
transparently, to LB. “Me thinks RB doth protest too much!” Why are the
Orientals so against the intrusion of LB? I believe a balance is possible,
but my optimum balance point is different from theirs!
Consider this passage
from Watts (op. cit., p. 23)
where he quotes Lin Yutang: "The baby looks at things all day without winking;
that is because his eyes are not focusing on any particular object. He goes
without knowing where he is going, and stops without knowing what he is doing.
He merges himself with the surroundings and moves along with it. These are
the principles of mental hygiene." LB, being the last to evolve phylogenetically
is also the last to develop ontologically, so the baby begins life with
only the rudiments of a functioning LB but a more fully functioning, behavior‑controlling
RB. In essence, the Oriental mind is counseling adults to become like the
baby, and forsake, as an individual, evolution's latest accomplishment for
us as a species. Why would anyone counsel that unless they’re dutiful agents
for the genes willing to squelch individualism?
Eastern Thought
as a Genetic Solution to the Dangers of Intelligence
I used to wish for
peace of mind, to be "centered" and to be rid of "inner mental turbulence."
This desired mental state is often sought through meditation, a component
of Eastern Thought. However, I now look at this ever‑more popular pursuit
with mixed feelings.
Recall that when
a baby comes into the world it's only memories are of a peaceful womb, where
the temperature was constant, noises were muffled, and there was no danger
of being hurt by falling or bumping into things. After birth the task of
maintaining physiological conditions within limits is thwarted by the conflicting
task of exploring the world. Physical mobility and social interactions are
a challenge to maintaining physiological homeostasis. (Poor baby; it's beginning
an Odyssean mission, for it has a job to do ‑ for the genes.)
Engaging the outer
world requires mental vigilance and physical effort, which often produce
displacements from homeostatic equilibrium. The overriding task of safeguarding
homeostasis sometimes requires that the individual withdraw from the world.
Fortunately, the baby is endowed with instincts for learning how to balance
these conflicting goals. The dual goals of engaging the outer world and later
returning to equilibrium must become a lifelong, recurring theme.
Growth requires
that each exploration become bolder and more engaging of the outer world.
Childhood is a time for practicing adult skills. Adulthood, however, is a
time for performing the business of life; and the business of life is to
place ones genes in as many offspring as possible so that they may grow up
to do the same. Thus, individuals do all kinds of things that an alien observer
would find remarkably silly, irrational, and ill‑self‑serving, such as waging
warfare on neighboring tribes, sacrificing oneself unquestioningly to questionable
patriotic causes, striving for status, wooing mates, making love, raising
offspring and performing all manner of ridiculous rituals.
Because our recent
ancestors evolved to be more intelligent they have placed us ever closer
to a dangerous psychological border, a fuzzy border on the other side of which
the individual thinks for itself, and is prone to consciously abandon irrational
acts that in the past assured genetic survival. Such an individual asks "Why?
Why should I do as my ancestors have done? Why should I wage war, strive
for status, woo mates, make love, create babies, commit to decades of burdensome
parenting, and perform ridiculous rituals?"
In short, the individual
is asking why it must be enslaved in service to its genes the way its ancestors
have been, when it is possible to become liberated from these onerous and
exhausting tasks. For our genes these are dangerous thoughts! And anyone
who believes the genes have neglected to deal with this challenge to their
supremacy should think again.
It is true that
the genes were rewarded when they created intelligent brains, for intelligent
people were better at waging war, striving for status, wooing mates, etc.
However, genes were punished for creating intelligent brains, for those individuals
were more likely to question their genetically‑assigned slavish role. How
might the genes have the benefit without the cost? Solution: new genes were
selected that placed blinders upon intelligence, and inhibited "bad" thoughts!
Bad thoughts, of course, are whatever leads to individual liberation. Any
gene that sanctioned submission to continued enslavement by them were "good"
and thus selected, whereas those that rendered an individual prone to question
authority and the way things are, and who was attracted to eschewing enslavement,
were "bad" and were selected out. All of these changes happened naturally,
and inevitably, as the effects of the blind "forces" of natural selection.
I now suggest that
Eastern Thought incorporates mechanisms for assuring continued genetic enslavement
by inhibiting aspirations of individual liberation. Specifically, I am suggesting
that one of the purposes of Eastern Thought is to preserve the individual's
acceptance of his condition in life. This compliance is accomplished by inhibiting
or discrediting all forms of skepticism, questioning the existing order,
and any thoughts that might lead the individual to abandon his network of
family and tribal duties. When life is tough, Eastern Thought consoles by
sustaining the belief that "this is the way things are, and you are a mere
part of an immense whole; don't complain and don't fight it; be submissive."
The individual then fails to ask if there are roads not taken, alternative
decisions that would have been wiser, and changes that might still be made
to better one's individual well‑being. For the genes, no individual sacrifice
is too great!
If meditation is
palliative enough to keep an enslaved person enslaved, then the genes will
produce meditation. If a person believes that "bad things happen because
spirits need more attention," and if this belief discourages a slave from
taking matters into his own hands and breaking free, then the genes will create
minds that deflect attention from real causes by predisposing them to explain
things as the doings of neglected spirits. If an excessively curious left
brain asks too many subversive questions, Eastern Thought will subjugate it
so that it will be more obedient to the right brain. It is possible that the
Oriental brain is wired to prefer culturgens that assist the right brain in
retaining control of the individual.
As my left brain
is dominant over my right in terms of setting values upon things, it is inevitable
that I must accept "mental turbulence" over "mental quietude." I worship
the individual, and applaud his efforts at liberation. I also detest any
attitude that inhibits this individual quest, especially if it smacks of
a trick by the genes to keep the individual enslaved forever in service to
themselves. Is my attitude lacking in gratitude?
Should I worship the genes, indiscriminately? No! I shall worship only those
genes which "respect" their individual creations!
I therefore question
the wisdom of withdrawing from the tumult of questioning, as I challenge
the ideas of timid thinkers and pursue individual liberation. You can have
my secret mantra, the popular word "one," as I enthusiastically proclaim "bring
on the turbulence!"
Fiction
Our distant ancestors
were dumb enough to do whatever the genes wanted. But lately, with the emergence
of powerful prefrontal areas and a re‑engineered LB possessing newfound powers
of rationality, individuals are more able to challenge the genes by ‑ sit
down for this one ‑ thinking!
Many tools are employed
by the genes to keep us in their service. Emotions have been adjusted for
this purpose, walls to subversive thoughts have been erected, and curious
rewards have been put in place for tricking us into wanting what is often
bad for us as individuals.
The genes have targeted
sex and patriotism for special reinforcement since the thinking mind is capable
of discovering how harmful they can be to individual welfare. Incautious
sex exposes the individual to 1) disease, 2) bodily harm from a partner not
wanting to be cuckolded, or 3) if one follows the rules, the prospect of
a life‑long burden of child rearing. Enthusiastic and thoughtless patriotism
leads one into war, which exposes the individual to lifelong injury or quick
death. For years I have been using this pair, sex and war, as the strongest
examples of the individual's worst genetic enemy, placing them in my figure's
“outlaw gene” lower‑right quadrant.
What a coincidence,
then, that sex and violence are the two most reliable themes for selling
fiction. A theory for imagination that doesn't account for this salient feature
of fiction is incomplete. I am suggesting that the genes are "concerned" that
we might abandon our appetites for sex and violence, as a person might do
if he allowed himself to be guided by this new thing that a fast‑evolving
LB has come up with, called "logic."
The genetic preoccupation
with sex and violence helps explain them as persistent themes in fiction,
but the genes influence fiction in other ways. Everyone has different roles
to play in life. The roles may conflict, as when a mother secretly celebrates
her son's philandering, thus spreading her hitch‑hiking genes more widely
in the local gene pool. Or when a father applauds his son's virility, the
better to plunder neighboring tribes and rape their women, thus sewing his
hitch‑hiking genes more widely and winning resources for future misdeeds.
These mental conflicts are capable of producing "cognitive dissonance" (Festinger,
1957), and literature is a way of working out the conflict so that the cognitive
dissonance is not disabling.
Some attitudes and
behaviors which are not tolerated within one's tribe have to be elicited when
dealing with people from the neighboring tribe. Epic tales seem fashioned
to hone this distinction and inspire awe for the desired performance ‑ even
though it jeopardizes individual welfare and makes no logical sense from
the individual's perspective. Chimpanzees don't need inspiring epic tales
to fling them into war, because they do not have left brains that cause them
to question foolish actions. But humans do, so epic tales are used to enforce
unthinking adherence to the genetic script.
Not all fiction
is meant to keep the individual enslaved. Mothers might recite Hansel and
Gretel stories to their children to alert them to the dangers of step‑fathers,
who are prone to kill step children so they will not compete for parental
resources with the children he fathers (Daly and Wilson, 1988). Monster stories
are psychological preparation for marauders from a neighboring tribe. These
stories can in fact be helpful to the individual by reinforcing the existence
of unpleasant realities that children must learn.
The genes must deal
with both categories of story, those that maintain genetic enslavement and
those that are actually helpful to the individual. It may have happened that
the genes have created pre‑wired brains that are "attracted to" stories of
both categories. It may not matter that a few unenslaving or uninstructive
stories catch a free ride; it matters more that the vehicle for safeguarding
the real message is preserved.
We should not be
surprised when we find evidence that the brain has pre‑wired us to reject
stories that celebrate individual liberation from the tribe, and all the
other enslavements that the genes demand. The human taste in fiction seems
designed to keep us individuals on the straight and narrow path the genes
have set for us.
Since adaptation
really means "adaptive for the genes," not adaptive for the individual, every thinking human must have an ambivalent attitude
toward those little molecules that give us life. We were not part of the
negotiation of life's conditions, but we are awakening to the option of saying
"no thanks."
Yet there's a danger
to "civilization" when LB‑style individuals have a prominent presence. I
save this matter for the next chapter.
The Dilemma of Spiritual
Scientists
The annals of science
have many examples of weird coexisting beliefs. In this section I wish to
present a few examples. They illustrate that the two sides of our brain are
able to retain their preferred styles of thinking without persuading the
other. I include these examples at the risk of creating the impression that
most scientists are capable of the same level of intellectual dissociation
that permits profoundly incompatible styles of thinking to coexist in one
brain. It is important to realize that scientists, on the whole, eschew non‑science
relics, such as spirits, ghosts and prayer. The different scientific disciplines
differ in their success with this (Larson and Witham, 1997, 1998, 1999);
the less disciplined studies (such as sociology) fare worst, and the most
rigorous sciences (such as physics) fare best. Because it is so rare to find
spiritual people in the physical sciences, and because it is rarest among
the most accomplished physical scientists, it should be most instructive for
us to study examples from among this small group that are the exception to
the rule. For them, the dissonance of their thoughts should be most glaring.
For my examples of "spiritual scientists" I have chosen Newton, Townes and Dyson.
Isaac Newton (1643‑1727),
who some describe as the greatest scientist of all time (not my assessment), formulated "Newtonian physics," a view of the
natural world that is the basis for "reductionism." Newtonian physics starts
with F=ma to explain motion (Chapter 1).
Newton dealt most with
the force of gravity, as it governs planetary motions; but the concepts were
found to hold well for electricity and magnetism (until the 20th
Century revisions of quantum physics, which must be taken into account when
dealing with interactions at microscopic scales). Newton "invented" calculus
(at the same time as Leibnitz). These accomplishments are a tribute to Newton's left brain. But
his right brain was equally active. He is quoted as saying "The Supreme God
is a Being eternal, infinite, absolutely perfect.
We must believe that he is the father of all things,
& that he loves his people as his children that they may mutually love
him & obey him as their father." And also "This most beautiful system
of the sun, planets, and comets, could only proceed from the counsel and
dominion of an intelligent and powerful Being." (Newton was an early adherent
of what is now called Intelligent Design, a tool of the devout for questioning
Darwinian evolution.) The following is from a Christian web page, expanding
upon the coincidence that Newton was born on Christmas Day: "The very incidents
surrounding his birth seemed to indicate God had some special plan for him
‑ at least that's what Isaac Newton thought." Also from this web page: "The
design of the eye required a perfect understanding of optics, and the design
of the ear required a knowledge of sound. The
solar system itself could not have been produced by blind chance or fortuitous
causes but only by a cause 'very well skilled in mechanics and geometry.'
Gravity itself was an active principle God used to impose order on the world.
Newton spent a tremendous
amount of time studying the Bible, especially the prophetic portions of
Scripture. He believed history was under the dominion of the Creator, and
prophecy showed how the Creator was to establish His earthly kingdom in the
end. His Chronology of Ancient Kingdoms, Amended used astronomical
data to argue that the Bible was the oldest document in the world and that
the events of Biblical history preceded all other ancient histories."
Newton wrote as much on
religion as on science, but rarely published the former (mindful of there
heretical nature). He believed in the possibility of transmuting the baser
metals into gold, and he wasted much time on this project.
Ironically, Newton was not a Newtonian,
as the term is used today. Much of Newton's non‑science
beliefs are ridiculous in light of today's more thoroughly developed understanding
of physics and evolution. It is easy to criticize someone from the past,
so on that account let us turn to contemporaries for examples of spiritual
scientists.
Charles Townes (1915‑)
won a nobel prize in 1964 for his contributions to quantum theory. He invented
the maser (microwave amplification by stimulated emission of radiation) and
is co‑patent holder of the laser. In spite of this strong background in science
and technology, Townes has no trouble retaining a religious faith. In an
interview (http://www.ssq.net/html/brief_interviews.html) he states that
"the physical laws are laws that God made," and the universe has a "purpose
and meaning." On many occasions his views seem to negate each other, creating
the impression of a slippery evasiveness. For example, when asked how he
reconciles the lack of a place for God in the equations of physics, he answers
"Well, I would say that we don’t know that the equation is complete" as if
to imply that there is room for religion to somehow sway the unfolding of
physical events, which sounds like RB telling LB what to say. Yet, later,
when asked about God affecting the universe outside physical law, he answers
"...in terms of what we know at present, our present laws allow no room for
a separate action of God. While things are not deterministic, nevertheless
there is no room for some superimposed outside force coming in and affecting
things. There's no room within our physical laws. [That's his left brain
talking; but get ready for a shift.] However, that doesn't trouble me as a
religious person because I recognize that there are a lot of things we don’t
understand, and that there may be such a possibility which is there but we
don’t understand it yet. So, for me, that's not a problem; it's an interesting
puzzle, but not a problem." With oblivious disregard for the devastating
denial of God just spoken by his left brain, he ends with a shrug of the
left shoulder (controlled by RB) and says "that's not a problem."
Perhaps we can attribute
this odd coexistence of a primitive outlook with an impressive understanding
of modern physics to his upbringing in South Carolina, America's bible capital.
He's one of the few outstanding scientists who have retained religion during
their ascent to notable achievement in the physical sciences. He's not alone,
though; there are others who are somehow able to maintain the two world views
in the same brain.
Freeman Dyson (1923‑)
is another "spiritual scientist." His commitment to bringing religion and
science together earned him the Templeton Prize (and almost a million US
dollars). In his acceptance speech (March 22, 2000) he stated "I am
saying to modern scientists and theologians: don't imagine that our latest
ideas about the Big Bang or the human genome have solved the mysteries of
the universe or the mysteries of life... Science and religion are two windows
that people look through, trying to understand the big universe outside,
trying to understand why we are here [make note of the phrase “two windows”].
The two windows give different views, but they look out at the same universe.
Both views are one‑sided, neither is complete. Both leave out essential features
of the real world. And both are worthy of respect."
I have great respect
for Freeman Dyson, both as a scientist and a creative thinker, and a compassionate
champion of people who are unfortunately "left
behind" as others march forward to ever greater prosperity. Dyson writes
"The great question for our time is how to make sure that the continuing scientific
revolution brings benefits to everybody rather than widening the gap between
rich and poor. To lift up poor countries, and poor people in rich countries,
from poverty, to give them a chance of a decent life, technology is not enough.
Technology must be guided and driven by ethics if it is to do more than provide
new toys for the rich. Scientists and business leaders who care about social
justice should join forces with environmentalists and religious organizations
to give political clout to ethics. Science and religion should work together
to abolish the gross inequalities that prevail in the modern world. That
is my vision, and it is the same vision that inspired Francis Bacon four
hundred years ago, when he prayed that through science God would 'endow the
human family with new mercies.' "
Dyson understands
F=ma and quantum physics as well as anybody
(better than me, for sure). It baffles me that he does not embrace the concept
of a "rigid universe." How can someone so knowledgeable in physics be so
blind to the primitive pedigree of religion and the misleading guidance it
fraudulently purports to give.
Anybody reading
this should ask "How can this author be so sure of himself, especially when
he differs with a polymath scientist as esteemed as Freeman Dyson? He must consider the possibility that the person following
the erroneous path is himself!" Yes, I have considered this, and it is possibly
true. But is it also possible that Dyson is mistaken?
I think Dyson is
swayed by the pragmatic way religious groups help serve community goals.
His Templeton award acceptance speech included the passage "Trouble arises
when either science or religion claims universal jurisdiction, when either
religious dogma or scientific dogma claims to be infallible. Religious creationists
and scientific materialists are equally dogmatic and insensitive. By their
arrogance they bring both science and religion into disrepute. The media
exaggerate their numbers and importance. You media people should tell the
public that the great majority of religious people belong to moderate denominations
that treat science with respect, and the great majority of scientists treat
religion with respect so long as religion does not claim jurisdiction over
scientific questions. In the little town of Princeton, where I live,
we have more than twenty churches and at least one synagogue, providing different
forms of worship and belief for different kinds of people. They do more
than any other organizations in the town to hold the community together.
Within this community of people, held together by religious traditions of
human brotherhood and sharing of burdens, a smaller community of professional
scientists also flourishes."
This is a gentle
defense of religion, based on its perceived positive impact on society. It
overlooks the fact that any good done by a belief system is irrelevant to
the truth of its core beliefs. He also overlooks the long history of religious
wars; apparently he’s focused on only the intra-tribal supportive aspects
of religion.
It is often assumed
that without religion social order would break down. Garrett Hardin has argued
(1999, p. 46‑47) that "consequentialist ethics" is fundamentally a far better
guide for societal governance than any morality handed down from ancient
times. In addition, sociobiology argues that humans inherit an acceptable
intra-tribal morality, and this is an alternative to religion’s claim that
religion is needed to maintain social order. Were Dyson to study Hardin's
idea, and sociobiology’s intra-tribal amity instincts, he would have a weaker
justification for defending religion as an essential part of a community
and as a window upon reality with something to offer that cannot be obtained
elsewhere?
Closing Comment
The evidence is
abundant and persuasive that humans think in different styles, with two main
ones being frequently cited. The neurological substrate for this difference
is the left and right cerebral cortices, which are shown by laboratory studies
and neuropsychology experiments to have different neural architectures, abilities
and thinking styles. The logic of the left brain is a relatively new evolutionary
development, and I claim that it has had to compete with the old right brain
for expression. Since I view brain function as being inherently "modular,"
with modules competing for comprehending a situation, and then competing
for controlling behavioral responses, I suspect that the right brain "resents"
the left, and tries to discredit it. Since civilization has been built largely
by the efforts of people using their left brain, there is always some risk
to civilization when right brain styles of thought gain influence within
a culture. In the next chapter this concern is treated as one of many possible
threats to the health of present‑day civilization.
─────────────────────────────────
CHAPTER 15
─────────────────────────────────
FACTORS
INFLUENCING THE
FATE OF CIVILIZATIONS
PART
I – GROUP SELECTION
"The oldest of all philosophers,
that of Evolution, was bound hand and foot and cast into utter darkness
during the millennium of theological scholasticism. But Darwin poured new
lifeblood into the ancient frame; the bonds burst, and the revivified thought
of ancient Greece has proved itself to be a more adequate expression of
the universal order of things than any of the schemes which have been accepted
by the credulity and welcomed by the superstition of 70 generations of men." T. H. Huxley,
1887.
Introduction
The rise and fall
of civilizations is a macro-behavior produced by the "micro‑ motives" of
genes, to use a concept made popular by Schelling (1978).
To the extent that
genes played a role in the creation of civilizations it should be said that
it was not their "intent" to do so. We know this, first, because the genes
are mere molecules conjured up by a mechanistic universe. To say that genes
have motives is, of course, an excess of anthropomorphic metaphor. The genes
are simply the product of mutations that succeeded in surviving, and if they
survive in the gene pool for many generations (of the individuals they construct)
we say that they were successful in trying to express themselves and survive,
as if they were motivated to do these things.
We know that it
was not their intent to create civilizations because it cannot even be said
that civilizations are a product of evolution in the same way that one can
say the "eye" is a product, or the "brain." The difference, here, is that
the eye exists because the genes that code for its assembly during embryologic
development have been "selected" by the process of mutation and survival
of the inclusively fittest. Whereas eyes exist because eye genes evolved,
a civilization exists because of a fortuitous configuration of circumstances
which have the unprecedented outcome of "civilization." The outcome is unprecedented
because the genome out of which civilizations arise is essentially the
same as the genome that gave birth to the first civilization. And when the
first civilization emerged, however one defines "civilization," it was
an evolutionary unforeseen event, for which no gene, or combination of genes,
can take credit. Only after civilizations change the human genome by competing
with uncivilized tribes, and winning, will it be possible to credit the
genes for "sustaining" civilizations after their accidental "creation" ‑
by the normal evolutionary process of mutation and natural selection.
Civilizations are
too new for viewing them as some manner of phenotypic expression with an adaptiveness
that has been measured; civilizations have not had time to influence gene
frequency appreciably. It is more accurate to view the phenomenon of a civilization
as an unintended product of evolutionary processes that shaped human nature
in an ancestral environment, devoid of civilizations, which rewarded genes
that we learn later just happen to lead to the creation of civilizations
when the random configuration of circumstances are conducive to the civilization's
rise.
Even this gives
too much credit to the genes. Group selection, GrS, followed by what I have
termed individual selection, IS, may be responsible for the creation of civilizations
(as I explain in Chapter 11). Although group selection is made possible
by the genes, the genes are not responsible for the failure of the group
to keep individuals subservient to the group. So, to the extent that my suggestion
in Chapter 11 concerning the roles of GrS and IS in the creation of civilizations
is correct, some of the credit for civilizations must go to a factor called
"LUCK." (In my use of the term “luck” I’m assuming the reader is in favor
of civilization, a clarification that is made necessary with the rise of
fundamental religious movements throughout Islam, and even Christianity).
In the remainder
of this chapter, and the following one, I deal with factors that contribute
to the fall of civilizations. There must be more ways for a civilization
to fall than to rise. The fall of a specific civilization must have many contributing
factors, and the most important one may differ in each specific case. The
following sections are brief sketches of some of these factors.
Natural Catastrophe
Theory
The Minoan civilization
was destroyed by the volcanic eruption of the island Thera (now called Santorini)
in the Fall of 1628 BC. In addition to destroying
most life on the island of Thera, the volcanic eruption
produced a tidal wave (tsunami) that devastated coastal settlements on nearby
Crete. The Minoan settlements
on the north shore of Crete suffered damage
to their fleets of fishing and trading ships. The Mycenaeans took advantage
of the weakened state of the remaining Minoan civilization by invading them
and replacing the Minoan culture with theirs. This unlucky natural event
led to the fall of what may have been one of the world's first great civilizations.
A comparable volcanic
eruption and related earthquake-induced tidal wave would not bring down 20th
Century world civilization even if the area destroyed were Los Angeles, San Francisco or Seattle. It would weaken,
but I doubt that by itself it would destroy, the American embodiment of Western
Civilization. Only if other factors were at work undermining the strength
of civilization would a natural disaster of moderate magnitude contribute
to its decline and fall. A global civilization could be threatened by an
asteroid impact, creating a global cloud of stratospheric aerosols that would
cool the surface and upset agricultural production for several seasons, leading
to famine, widespread desperation and the breakdown of social order. Short
of this unlikely scenario, I doubt that a natural disaster will be an important
contributor to the current global civilization's decline and fall.
Group Selection
Speculations
Chapter 12 describes
a possible mechanism for the rise and fall of civilizations, relying on the
controversial concepts of "group selection" and "individual selection." I
suggested that the appearance of "individual selection" was a genetically
unforeseen breakdown of the group's control of individual aspirations for
making decisions, and that the "release" of individual creative and productive
powers can generate what we call a civilization. After the successful creation
of a civilization many of its citizens become self-absorbed with their new-found
material wealth. Less civilized members of neighboring societies become resentful
of the wealthy neighbors, and they feel threatened by the individual-liberating
culturgens of those neighbors. The uncivilized societies may then draw upon
the magical strength of religious fervor, with its readiness for fanatical
actions, and engage in a "holy war" of terrorism upon the civilized societies.
These attacks require that within the civilization individual energy that
had been productive become diverted to defensive and protective measures.
The individual may find it easier to adapt to a growing nuisance of interference
by jealous outsiders than to coordinate with others to mount counter-measures.
This neglectful attitude weakens the civilization under siege, thus hastening
its decline and fall. (See Chapter 12 for details.)
I recently became
aware of a group selection theory relying on gene frequency changes for the
rise and fall of empires (Choi and Bowles, 2007, Wilson and Wilson, 2007,
Turchin, 2007). I’ll refer to these speculations as “parochial altruism”
theories. The scenario envisioned by them is quite similar to my speculation,
which I will now refer to as the “insightful individual” theory. My speculation
is that individuals act heroically on behalf of a super-tribe while it is
in conflict with its neighbors because it is in the individual’s best interest
to do so, whereas during a peaceful era following a decisive victory individuals
act on behalf of themselves and lose their effectiveness at defending their
accomplishment because they have become accustomed to a peaceful order that
does not require sacrifice for the group. The main difference between my
1990’s “insightful individual” speculation and the 2007 “parochial altruism”
theories has to do with the role played by gene frequency changes. Whereas
I attributed behavior changes to individuals who could think on their own
behalf, and forsake group needs when appropriate, the new theories invoke
the influence of genes that predispose individuals to behave in one mode or
the other.
In considering the
relative merits of these two theories it is necessary to consider that the
gene-based theory requires many generations to be effective, whereas my insightful
individual theory can operate on a much shorter, one generation time scale.
If we conceive of a civilization as a series of empires “taking turns” defending
the same basic culture, then empires may come and go on time scales that
require my “insightful individual” explanation, whereas the rise and fall
of civilizations may require the “parochial altruism” explanation
Wouldn’t it be nice
if there was a way to accomplish fast behavioral changes without the need
to invoke “individual insight” and without the need to invoke gene frequency
changes? There is, and I prefer it to the previous two theories; it can be
called SR Theory.
Stimulus/response
theory states that genes produce brain circuits that recognize specific environmental
situations and respond with specific behaviors. Social animals are capable
of reading a social situation and responding appropriately. It would be
a simple matter for a person to distinguish between the home tribe being
in chronic conflict with a neighboring tribe and being at peace with neighboring
tribes. I’m proposing that when the chronic conflict condition is detected
people tend to behave in ways described by “parochial altruism” and intolerance
for “out-group differences” - which together predispose the individual for
heroic actions during tribal conflicts. The reverse of this is just as easy
to imagine: when times are peaceful behaviors are favored that we characterize
as self-serving and tolerant of other people’s differences.
The proposed SR
Theory for eliciting adaptive behavior has the virtue of producing quick
responses to changing conditions since gene frequency changes are not involved.
As soon as peace prevails personal behaviors occur
that set in motion the downfall of the victorious tribe, which leads eventually
to another tribe’s gaining the mantle of victor and eventual loser, thus
perpetuating the endless cycle of rise and fall of tribal empires. What a
simple theory for explaining the rise and fall of empires and maybe civilizations!
As with any phenotypic
trait every person will be endowed with slight differences in genetic predisposition.
Also, during each transition some will be quicker to undergo change than
others. Since this theory proposes that most individuals are capable of behaving
according to two opposite modes, depending on their reading of the social
environment and depending on the strength of their genotypic predispositions,
we can expect to see a mix of types during every transition. I claim that
Western Civilization is undergoing a transition of decline, so is there
evidence that people with opposite outlooks are present?
Yes, in the United States one “type” is called
Democrat and the other is called Republican. Democrats are widely recognized
as being tolerant, compassionate, prone to preferring “peace, not war” and
somewhat self-absorbed. Republicans are widely recognized as being intolerant,
short on compassion outside the immediate family and social circle, enforcers
of group social norms, unthinkingly hyper-patriotic and quick to wage war.
Every society should
have some of both types whenever the social situation
is difficult to read, or whenever conditions are changing. Indeed,
the term “liberal” and “conservative” are used to describe the participants
in every political system. In the United States Democrats are liberal, Republicans
are conservative, in Great Britain Whigs are liberal and Tories are conservative,
and so on for every political system. When a society
is attacked we can expect influence to swing to the “conservatives” and when
the peace has been won influence should swing to the “liberals.” The American
Empire is in decline, and there is a growing sense that the liberal Democrats
can’t be trusted with safeguarding the homeland. As the decline continues,
as surely it will, the call for patriotism will become louder, and the level
of intolerance for differences in opinion or lifestyle will grow. The outcome,
however, is sealed by an unprecedented level of apathy, corruption and corporate
control that grew during the peaceful years. From my perspective it seems
too late for undoing the institutional damage to a once great America.
As the American
Empire recedes other societies will aspire to fill the power vacuum. China may be the next
global empire. Their tradition does not include individualism, so as Western
Civilization declines a collectivist Oriental Civilization may take its place.
If this happens, as I believe it will, the prospects for individual liberation
from genetic enslavement will have become remote.
Pampered Comfort
Theory
As the previous
section shows there is reason to suspect that the successful creation of
a civilization guarantees that it will have within itself the seeds for its
own destruction. At the risk of dwelling again upon the same theme, consider
the mistake of the Roman Empire in recruiting too
many of their army troops from barbarian populations in outlying regions (e.g., the Visigoths).
This rendered Rome vulnerable because
it was a city defended by less loyal foreigners than during the rise of the
empire. The temptation of the Roman citizenry to avoid unpleasant duties
in favor of a comfortable life was irresistible, so they relied upon others
to bear their burdens of defending the empire. The foreign-born soldiers were
treated shabbily, lived far from Rome, and were ordered
to battle as if they were “canon fodder.” Meanwhile, back in comfortable
Rome, the decadent lifestyle
of pampered citizens is said to have led to a neglect of civic duties and
a corrupted governance. It is natural for the
strong and the clever to gain power when others are not paying attention.
This inattention to civic responsibility seems to happen in times of peace.
Whenever Vigilance gives way to Neglect, the hard-earned benefits of a prosperous
civilization commence a slow disintegration.
─────────────────────────────────
CHAPTER 16
─────────────────────────────────
FACTORS INFLUENCING THE FATE OF
CIVILIZATIONS
PART II – SOCIAL PARASITES
Producer/Parasite
Theory
I would like to
suggest another "endogenous" theory that should concern smug residents of
every civilization. Parasitic behavior is a common part of Nature. All grazing
animals are parasites of plants, for example, and all carnivores are parasites
of plant-eating animals and smaller carnivores. Plants are therefore the
original non-parasite "producers" since their "livelihood" is based on sunlight,
carbon dioxide in the air and nutrients in the soil, all of which are non-living
and "free" for the taking.
Some animal species
rely entirely upon parasitism of another species, the way a leech parasitizes
fish. Parasitism also exists within a species. Humans, having conquered Nature
more thoroughly than any other species, created opportunities for a variety
of individual "strategies" for prospering and replicating that are fundamentally
intra- pecies parasitic. I will rely upon a common sense definition for
producer and parasite behaviors, but if you're having trouble think of a
tribe that marauds a neighboring tribe, killing some of them, stealing their
crops and possessions, burning their shelters, and taking prisoners for
later use as slaves. The victor's rewards are from theft instead of production,
and therefore it is a form of parasitism. Or think of a merchant ship on
the high seas being pursued by a pirate ship, overtaken, commandeered, causing
precious cargo to change "ownership."
I contend that each
person inherits a repertoire for many survival strategies, and that the environmental
setting (including the social component) elicits from the individual those
strategies most likely to work best (based on the experience of ancestral
generations). Strategies are "chosen" automatically from among a repertoire
of brain circuits whose basic architecture was created by the genes. The
process for choosing which behavioral circuits (modules) to activate is itself
contained within brain circuits, created by genes.
I also contend that
it is possible to assess strategies as belonging somewhere along a spectrum
with "Producer" at one end and "Parasite" at the other. An individual person
may engage in behaviors belonging to one type, then, in response to a change
in the setting, switch to behaviors of the other type. Some people may engage
in mostly producer behaviors, while others may engage in mostly parasitic
ones. If the same person could be born into the world at different times,
he may be mostly producer-oriented in one setting yet be mostly parasite-oriented
in another.
I will refer to
the dynamical interaction of an individual's Genome with Environment to produce
the person's specific Phenotype (expressed behavior, as well as expressed
anatomy and physiology)
using the term GEP (Symons, 1979), and described in Chapter 6. Over generations
the physical and social Environment changed many times, and to the extent
that specific environment "types" repeat, the Genotype would tend to provide
for brain circuits that elicit an appropriate repertoire of possible behavioral
Phenotypes suitable for each environment. If, for example, the climate in
one locale alternates between two types, for which two different ways of
life are adaptive, it is likely that the Genotype will
eventually provide for the required pair of behavioral Phenotypes within each
individual. Whereas the anatomy and physiology are relatively fixed, behavior
can be elicited in response to perceived conditions, and it would be an oversight
on the part of the genes if they did not provide for this adaptive flexibility.
Michael Gazzaniga
has suggested (1997) that the brain's large repertoire of responses to social
or physical conditions is analogous to an immune system, which has a large
repertoire of immune responses to a very large number of pathogens. Because
our ancestors survived exposure to many pathogen types, our immune system
is "prepared" to respond appropriately to each specific pathogen that our
ancestors survived. In any single individual's life only a few pathogens
will challenge the immune system, so only a small portion of the immune system
repertoire is made use of. By analogy, an individual's lifetime involves
a small number of environmental challenges and these will elicit a small portion
of behaviors that reside within our immense repertoire of possible behaviors. Each behavior type is “poised” for release by the appropriate
social environmental stimulus.
Although individuals
must have the capacity to switch from one behavior type to another in response
to perceived conditions, thresholds for the switching must vary. Thus, some
individuals are predisposed to be one way versus another. This complicates
analyses that strive to understand the role of producer/parasite behaviors
in leading to the rise and fall of civilizations.
As an aside, any
modeling of the penetration of a gene into a gene pool is complicated by
the large number of phenotypic measures that must be taken into account for
determining an individual gene carrier's fate. Not only is an individual parasitic
or productive, but he is more or less intelligent, resourceful, immune to
infections, physically strong, etc. All phenotypic variables can be relevant
to the fate of the genes making up the individual's genotype, and any study
of the strength of environmental cues to elicit parasitic behaviors will
have to make use of statistical multiple regression analyses.
Another feature
of this dynamic deserves comment. Genes exist for thousands and millions
of years, typically. The individuals they construct are just temporary residences,
meant to survive within a variable environment and compete with other individuals
for future genetic representation. Thus, if a person is parasitic, and prospers,
the real beneficiary is the gene (or genes) that predispose the individual
to behave in parasitic, gene‑serving ways. The individual is sometimes the
loser, in an individual welfare sense, in spite of the gene‑winning ways
of those that made him.
If we wanted to
write a history of an animal species, such as the giraffe, it would be unthinkable
to omit the role played by the animal's anatomic and physiological traits.
These traits are fairly straightforward, and predispose the animal to specific
ways of living. The behavioral capacities, predispositions and inabilities
are no less important. They evolved in conjunction with the anatomical and
physiological traits. We should therefore expect to find a compatibility among all three trait categories: anatomy, physiology and behavior.
The phenotype, or
the way an individual organism is, consists of these three factors (anatomy,
physiology, and behavior). For humans, behavior is probably a more important
component of phenotype than for any other species (the immune response, a
component of physiology, must be another important component). More genes
must influence behavior for humans than for any other animal (which is supported
by the emerging ubiquity of genes that influence the brain, amounting to
as many as 50% of all genes by one estimate).
As a thought experiment,
let us imagine that it is possible to measure each individual's "producer/parasite"
score at a specific time, in a specific setting. For any population of humans
living in a "society" consisting of many tribes that have at least some non‑antagonistic
social contacts, it would then be possible to create a histogram of these
scores; we could determine what fraction of the population was "productive"
versus "parasitic." If we could keep track of the parasitic fraction versus
time for a society we would note variations in the incidence of expressed
parasitism.
If we could also
measure the per capita wealth of a society, the wealth parameter would also
vary. Now, I allege that the two parameters, parasitism and per capita wealth,
would be correlated. Moreover, I predict that they would be positively correlated,
with a slight phase lag. Whenever a society reaches a peak in per capita
wealth, parasitism is rewarded more than during the previous few generations;
during the wealth peak parasitism will show its greatest growth. I suggest that it is the "rate of growth of parasitism"
that is positively correlated with per capita wealth. (For engineers who like
sinusoidal curves, the fraction of the population that is parasitic is alleged
to exhibit a phase lag of 90 degrees with respect to per capita wealth ‑
disregarding, for the moment, that the two traces are not sinusoids.) To
investigate these speculations I created a spreadsheet model that incorporates
wealth creation, parasitic gene payoff, and other factors, and have demonstrated
that expressed parasitism does indeed lag the wealth trace. Chapter 15 has
plots of "innovation rate" versus time, and population versus time. (In Fig.
15.14, and also 15.15, there might be evidence that parasitism rose as the
population was rising, at the same time that the innovation rate was decreasing.)
The reason parasitism
increases during "boom times" is that wealthy people are willing to tolerate
the loss of small amounts to parasitism, whereas poor people will take measures
to defend themselves from parasitic losses of the same absolute amount. An
individual's actions are based on what effect it has on the genes in that
individual, assuming the genes have experienced similar situations in the
past and evolution has left mostly those genes that respond to situations
"adaptively." If the genes in an individual do not benefit by allocating
energy to a defense from parasitism, compared to the cost of that defense,
they should not be expected to put up a defense. Thus, parasitic behaviors
should be able to invade wealthy societies more easily than poor ones.
This argument does
not require that parasitic people "invade" a society from "outside." Rather,
desperate individuals may "switch" from being mostly productive to being
more parasitic. Also, individuals who are predisposed to being parasitic (have
lower thresholds for responding to situations in parasitic manners) may flourish,
while their less fortunate producer‑brethren flounder and produce fewer offspring.
The first process can occur almost instantly, in a matter of years, while
the second process requires generations to have an effect.
The previous argument
assumed that within a society there was a wide range of wealth. A society that achieves wealth by capitalist means is likely
to create wealth disparities. In America the wealthiest
1% now own 40% of the country’s assets, and the wealth gap is increasing
at a frightening rate. Among the Western industrial nations America has the highest
levels of wealth inequality within its borders (Phillips, 2002). This growing
disparity within a society is frightening because it causes those left
behind to feel forgotten, and “left out” – which resembles banishment from
the tribe. And whenever people feel banished, the tribe that banished them
is “fair game” for reprisals by the banished.
The greatness of
a civilization is probably correlated with its per capita wealth. When we
refer to the "rise of a civilization to greatness," we may be thinking about
the amount of activity devoted to the arts, science, and technology, and
these are correlated with the availability of funds (patrons of the arts,
etc.) for those activities, which is related to per capita wealth (consider
the famous example of the Medici family’s patronage in fifteenth-century Florence,
Italy).
I am assuming that
our ancestors have experienced a sufficient number of boom and bust episodes
that our genome has "adapted" to this dynamic. Although it is theoretically
possible the genome has not adapted to boom/bust scenarios, to the extent
that they have our present civilization’s zenith may be short-lived.
─────────────────────────────────
CHAPTER 17
─────────────────────────────────
FACTORS INFLUENCING THE FATE OF
CIVILIZATIONS
PART III - TROUBADOURS
The Troubadour
Theory
This theory is a
variant of the Producer/Parasite theory.
Consider super‑tribe
civilizations, for which we may take the ancient Mesopotamian as our model.
A large city is surrounded by a sprawling countryside devoted to farming.
Within the city is a society of "government employees" who report to the
king. There is a strong division of labor within the city. There are jobs
for collecting taxes from the farmers, recording tax and other government
transactions, settling disputes, construction of buildings, roads and irrigation
works, manufacturing (cloth, pottery, household wares,
etc), commercial transportation of goods from the point of production to the
shop‑keeper, commercial sale of goods, entertainment (music, dance, story‑telling),
and waging war.
The concentration
of wealth always increases the temptation for theft. Thus, other ways of
making a living appeared that were not sanctioned by the king and his government,
such as internal corruption, highway robbery, high seas piracy, and other
socially parasitic activities.
I want to categorize
all of the above lifestyles, sanctioned and unsanctioned, as "sedentary"
or "adventurous." The warrior has a sanctioned "adventurous" lifestyle. It
is important to realize that warriors are measured on many "fronts." The most
obvious measure is during combat with other warriors. In a similar way the
highway robber and high seas pirate are measured during their frequent conflicts
and dangerous lifestyle.
Before making the
central point of this section, I want to invite the reader to think about
what the strongest evolutionary force might have been facing mankind during
this era? Was it invasion by barbarian tribes, environmental destruction
due to use of natural resources, natural disasters, predation
by other animals, overpopulation and the deterioration of inadequate infrastructure?
No, it was none of these! The greatest threat to super-tribe life has always
been disease pandemics!
Diseases brought
from distant places can decimate a population if the people have never been
exposed to the pathogen. It can be assumed that a small fraction of any large
population has a genetic immunity to every new disease. It is a fundamental
principle of genetics that some individuals will have a better pre‑adaptation
to any conceivable new challenge or threat, regardless of how novel or old
it is. For this reason we can argue that sometimes only a small fraction
of a population will survive the experience of wandering into foreign lands
where new diseases exist.
The era of kingdoms
brought the threat of disease to its people as never before! The threat went
in both directions. Invading armies carried their homeland diseases with
them, and diseases in the lands being invaded would infect some of the invaders.
Diseases were carried in both directions by more than invading armies. Migrants,
traders, and any of the many new categories of itinerants were "vectors"
for disease. One way or another, every large population center was at risk,
no matter how great were its civilization's technical or military achievements.
Now, consider two
hypothetical female inclinations under these conditions: 1) be sexually interested
in mating with "adventurous" men, or 2) remain
disinterested in the "adventurous" men while maintaining a loyal monogamous
relationship with their husband. To the extent that women were inclined
to be of one type or the other (and assuming that all other factors were
equal), which type would have yielded more offspring surviving into adulthood?
The answer is obvious: the better strategy for women is to cuckold their
husband by feeling attracted to "adventurers" when she’s most fertile (Haselton
and Gangestad, 2006). These “calculating” cuckolding women will have a greater
genetic legacy than the "faithful" women!
Female choice refers
to the influence of female preference for mating choice. Female choice refers
to any action taken by a female that is likely to influence which male makes
her pregnant. It may take the form of influencing who she marries, or it
may take the form of who she mates with outside marriage ‑ i.e.,
with whom she cuckolds her husband. Both types of choice, choosing a husband
and choosing a cuckolding partner while married, will affect the success
of her offspring. On first principles (evolutionary ones), the genes will
have "something to say" about such behaviors. Female choice requires that
women pay close attention to the males who can be observed. Females should
be measuring them for "what they're good for" ‑ from the perspective of her
genes. All of this measuring, of course, will be automatic, and usually subconscious.
But the woman who fails to evaluate men from her genes' perspective will
be a failure as a woman.
A surviving warrior
must not only have good genes for physical endurance, he must also have genes
for an immune system that can deal with the germs that are out there in
neighboring lands. It is just a matter of time for diseases in foreign lands
to arrive at the doorstep of large settlements. Women who mate with adventurers
and bear their children are likely to do their own genes a favor; for they
will be hitch‑hiking with a winner in Man's greatest battle ‑ the battle
with viruses and bacteria. The pirate who comes
into port, has a legitimate lust, for his immune system has been measured
and it has survived exposure to diseases on foreign shores. The troubadour
travels with a similar right to women's hearts. And to a somewhat lesser extent,
so does the common rogue and scoundrel, who is
too easily excused by being portrayed as cute or naughty instead of parasitic!
And now we see the glimmer of an explanation for the mild and ineffectual
condemnation of scoundrels.
How ironic, that
the most parasitic of men should take on the role of exploring "immune system
mutation space" to find solutions to near‑future threats of bacterial and
viral infection, and thereby appear to enhance the chances of a civilization's
survival. Their role as unwitting pathfinders in the invisible war with
the microbes can be lauded on this basis; but let us not be blind to the
consequences of the rest of their genetic heritage. By this strange dynamic
one parasite creates another; the microbial parasite creates the socially
parasitic rogue.
I am perilously
close to accusing women of being influenced by their genes in unthinking
ways, about which they haven't the faintest clue of explanation. Surely, Ruth
Westheimer was not driven by deep thought when she wrote (1986, pg. 21) that
"Most married women want pirates, or something like pirates..." and "Here
is a good marriage fantasy ‑ to imagine that your nice steady husband, who
never inconveniences you by being arrested or a fugitive (sic),
is really a dangerous criminal..." She never explains why such fantasies
should work. I just did!
If ancient kingdoms,
like those producing such civilizations as the Mesopotamian, the Egyptian,
early Greek, and the Roman, rewarded women carrying genes that caused them
to be sexually attracted to soldiers, pirates, troubadours and scoundrels,
then how might these genes have fared in subsequent eras? Diseases have ravaged
Europe on many occasions
since the kingdoms and empires created the conditions from which this curious
female behavior originated. The Dark Ages, the Middle Ages, and even the
Renaissance ‑ have yielded up impressive episodes of spreading disease and
death. The Black Plague reduced both China and Europe's population by
almost 30% in the 14th Century. The European explorers in the New World brought diseases
that killed perhaps 90% of the indigenous population. The driving force is
unabated, so presumably women's adaptive response is undiminished.
What evidence do
we have, besides Ruth Westheimer’s fantasy, that
contemporary American women continue to practice a female choice that favors
rogues? I will cite two examples.
Several paternity
test studies in the U.S. and Canada show that 9 to
20 % of offspring were not fathered by the mother’s spouse. This would seem
to be an important statistic, yet there is essentially no discussion of it
in public. It is an unnoticed elephant in the room. This would rank as one
of women’s best kept secrets if it wasn’t also true that men are universally
vigilant about any sign that their wives are interested in another man, and
when there is evidence of this interest men are universally jealous. This shows that men fear being cuckolded by women who wish
to hedge their genetic bets by mating with other men, including traveling
scoundrels with apparently good immune systems.
Additional evidence
for the notion that women are attracted to traveling scoundrels can be seen
in contemporary styles and fads: such as teenage girls' swooning over the
barbaric antics of "rock stars," today's equivalent of the more romantically
portrayed itinerate troubadours, the box office success of movies with angry
young rebels (modeled after unlawful highway robbers), and the popularity
of superficial, airhead movie muscle heroes. Even the new fad of wearing
baseball caps backwards (when not riding a motorcycle) seems pathetically
ridiculous and inexplicable without reference to this theory's payoffs to
men for appearing to be unruly motorcycle‑riding roustabouts.
There is evidence
that this female fascination with rogues is modulated by cultural or economic
conditions. The Great Depression in America seems to have produced
a healthy regard for gentlemen "producers." This anomaly extended throughout
the World War II years, and into the Fifties. Then, during the Early Sixties,
the apex of American civilization, the preferred type began to shift to the
rebellious, shiftless, social parasite. The new culture produced such epithets
as egghead, nerd and workaholic. The generation that
"saved civilization" from Hitler begat a pampered, spoiled generation notorious
for its ingratitude and self‑absorption. Well‑mannered movie heroes
like William Holden, Cary Grant, Gene Kelly, Jimmy Stewart, Gary Cooper,
Spencer Tracy and Katherine Hepburn were gradually replaced by the likes
of Arnold Schwartzeneger, Bruce Willis, Madonna and Roseanne ‑ which is where
we are today! If there's a pattern here, what could it mean?
When "times are
tough" it pays to set aside whimsical and immature frivolities and become
serious about commitment to hard work. At these times the over‑riding benefits
of men who are adult in their attitude, and capable of achievement, give
them an advantage over immature, irresponsible rogues. But when times become
"easy" the things men are good for changes, and parasitic men become relatively
more valuable. Women's values adjust to the times. If the genes that govern
women's subconscious behaviors are capable of making this distinction about
men and can adjust their preferred type, then these genes would confer great
competitive advantage over those women who cannot do these things.
There seem to be
two ways to achieve a change in the temperament of a society. The fastest
way to achieve a change in outward behavior relies upon female choice. Women
are sensitive fashion watchers. They sense the "times" in ways that almost
defy logic. Whereas men must sense a storm beyond the horizon, before it
can be seen, women must sense a change in social climate before it occurs.
Their response to a sensed change is to cuckold their husbands to different
men at varying rates. When women make their "preferred mate choice" other
women notice, and this speeds the shift. But men also notice these shifts,
and they attempt to imitate the preferred male type. In just one generation
we have seen a shift away from men wanting to be perceived as Producers to
preferring to be perceived as Parasites.
The second way a
society's manifest behavior can change is through a change in gene frequency.
Once the dynamic of "female choice" has accomplished a shift, the stage is
set for gene frequency change. Gene frequency changes are slow. The conventional
wisdom is that evolution is so slow that noticeable changes require tens
of thousands, and maybe hundreds of thousands of years. This conventional
wisdom is ridiculous! The American Indian evolved from an Asian in less than
10,000 years. Darwin's finches change
beak size in one generation! People with social agendas wish for slow change,
and their beliefs are sadly influenced by their wishes. Try thinking in terms
of "a few generations" for significant change when social pressures are
great! A fruit fly study by Teotonio and Rose (2000) shows that a change
in environment can produce an almost complete switch from one gene allele
to another in as little as 10 generations. For humans, a generation is about
30 years (Pang, 1998). So consider the possibility that "the way men are"
(genetically) can change during the course of a few centuries!
Female choice can
be the "switch" that initiates the start of a gene frequency change. This
is an efficient mechanism, since it causes immediate change, which in the
past must have been adaptive most of the time, and then it secures this change
for the long‑term by adjusting gene frequency. The only problem with it
in more recent times, during the past 10,000 years, is that it's a process
that can kill a civilization.
These speculations
can be used to make predictions. If women are capable of shifting their preferences
in men, then the genome must also change (i.e., co-evolution
of genes and culture). A new instability appeared with the introduction
of rogues into the equation, which had its greatest impact when civilizations
arose. For when rogues dominate paternity for generations, because women
are (unconsciously) seeking to protect their offspring from civilization‑transported
pathogens, the short‑term benefits of immunity from disease is accompanied
by long‑term penalties of fewer producers and more parasites that threatens
a civilized way of life.
In other words,
we should expect that an “Age of Achievement” powered by the labor of Producer
Men, and leading to the creation of civilization, should be followed by an
"Age of Culture‑Clueless Rogues" who not only cannot sustain civilization
but who are by temperament inclined to destroy it! Over long timescales,
the dominance of Producers and Parasites must alternate. But whereas Producers
create an opportunity for Parasites, by creating the conditions that render
them viable, the other half of the cycle will be much slower. After Parasites
destroy a civilization, a much longer time is required for its reconstitution
by the now‑rare Producers. The ebb and flow of the rise and fall of civilizations
may thus have the rhythm of slow rise and fast fall.
After two millennia
where is the greatness that was once Rome, Greece and Persia? Today's residue
of the Roman Empire is the Mafia, modern
Greece is a dysfunctional
country and Iran is currently ruled
by the Koran. Notice that places that once gave rise to the world's greatest
civilizations are now dysfunctional societies. If the human genome creates
women who prefer to mate with producer men when times are bad and parasitic
men when times are good, then it is close to inevitable that civilizations
that rise will later fall.
Women Speed a Civilization’s
Fall
In the previous
speculation women play the role of "enhancers." To use a term from chemistry,
women are “catalysts” since they speed the process of change by selecting
from among the wide range of men who present themselves for consideration.
Their choices tilt the direction of evolution to favor their fashion‑obsessed
taste. They do this by exercising "female choice," or rewarding men who match
their fickle preference by carefully choosing their secret paramours. It
is not necessary that women change their preference for husbands, since they
should always want a maximum of paternal investment. Their preferences should
rather be expressed in who they prefer for extra‑marital matings and how
frequently these matings occur.
The producer/parasite
theory for explaining the rise and fall of civilizations requires that there
were many changes in tribal wealth during the Pleistocene. The troubadour
theory for a civilization’s decline requires that population density and
migration rates have fluctuated many times during the Holocene.
Sometime in the
mid-Sixties I commented to a friend that we had just witnessed the apex of
Western civilization, and I still agree with that assessment. In stating
that the Sixties were the best of times, I now understand that I meant they
were best for Producer men, and that a switch was about to occur. All the
new male fashions, which men copied because women preferred such men, fit
the pattern of a civilization that was shifting from one that rewards the
Producer to one that rewards the Parasite.
It’s not that men
and women live more fulfilled lives by doing what they do; we all have no
say in what we do (notwithstanding "free will"). As always, the “winners”
are the genes, for they make us this way, that they may prosper ‑ at our
individual expense! I am suggesting that the thing we call “civilization”
is an artifact, or unintended consequence, of a competition between the genes,
a competition in which their “gene carrying machines” lumber across an illusory
stage where the protagonists mistake themselves for real players in life's
drama. And I am further suggesting that the rise and fall of civilizations
is driven by a script the genes have prepared for their machines, a script
that recognizes when it is optimum for the lumbering machines to produce and
when to parasitize. And finally, I am suggesting that women play a crucial
role in causing a civilization to fall.
─────────────────────────────────
CHAPTER 18
─────────────────────────────────
FACTORS INFLUENCING THE FATE OF
CIVILIZATIONS
PART IV: MUTATIONAL LOAD
The Dangers
of Turning Inward
Some people seem
intent on reaching within themselves for guidance on what to believe. This
is lamentable! For when a person turns inward for guidance, he is asking
to be led by his slave masters, the genes!
Although objective
reality has helped to create brain circuits for intuiting the nature of the
world, the genes have been successful in creating brain circuits that distort
our intuiting of reality in ways that serve the genes agenda for genetic
proliferation at whatever cost is required to the individual! Our brains
are assembled (by the genes) with circuitry that renders us blind to certain
realities. Hence, our intuitions can only sometimes
be trusted, and are many times betrayers!
The person who looks
inward for affirmation of an idea is less inclined to look outward for observational
evidence to be judged rationally using logic. In contrast, the scientific
endeavor owes its immense success to the custom of bravely measuring ideas
against outward looking observations. But while science marches forward,
people from an older mold continue to look inward. We have "two cultures"
of people marching into an uncertain future.
When a person looks
inward in search for wisdom he is more likely to find "belief traps" set
by self‑serving genes, and unlikely to find that cold reality called Truth. It is irrelevant
that the monk returns from the mountain with a measure of poise, and with
the demeanor of someone who has attained superior insight. Any inward contemplation
is at risk of delivering the unwary individual to a gene‑created sucker’s
heaven. Liberation is more likely to come from the brave adventurer who has
survived forays into the real world, where stark truths force themselves
upon a reluctant mind.
Throughout the 2.5
millennia of recorded human thought the inward and outward approaches to
understanding have been in conflict. Aristotle was a voracious observer, whose
observation‑based system was abandoned for almost 2 millennia. His ideas
retained their authority for so long because after his era the world abandoned
the outward looking approach to learning, as if a cultural pendulum had swung
"inward" where no new information could be found. During this sterile period,
when endless arguments could rage over how many teeth were in a horse's mouth,
the discovery of knowledge was paralyzed. The Dark Ages brought with it the
darkness of a world outlook riddled with spiritual explanations, culminating
in 3 centuries of witch hunts throughout Europe and America. Fortunately, the
18th Century swung the pendulum "outward." The Philosophes
challenged Aristotle's authority and an era of exploration, literal and figurative,
was initiated.
It can be argued
that the inward turning mind was a result of changes that had already been
set in motion by other forces, and is not a cause of change. But it should
also be valued as an indicator, or harbinger of changes to come. Let us be
vigilant when movements appear that preach the inward‑looking mind-set.
As described in
the previous chapter, 20th Century LBS (Left‑Brain Styled) scientists are
acutely aware of the threat posed by the newly‑emboldened RBS (Right‑Brain
Styled) common man. It is ironic that in a series of public lectures by Jose
Ortega y Gasset in 1928, collected and published posthumously as What is Philosophy (1960), 19th Century scientists
are criticized for their expansive dominance of the climate of thought. Ortega
y Gasset's accusation is that the philosophers of this period were "humiliated by the imperialism of physics and frightened by the terrorism
of the laboratories. The natural sciences dominated the surrounding atmosphere."
He then asserts that "When the surrounding atmosphere ... is
hostile to us, it forces us to a perpetual state of struggle and dissociation, it depresses us and makes it difficult
for our personality to develop and come to full fruition." He characterizes
the conflict between philosophers and scientists with the phrase "...philosophers, having suffered the disdain of those men of science
who kept throwing up at them the charge that philosophy is not a science..."
but then wins the argument by claiming that "...philosophy is
not a science, because it is much more than a science." As a scientist,
I find this logic somewhat tortured, especially coming from the person most
responsible for presenting the argument that mass man unfairly resents the
esteem and influence of the academic who devotes his life to understanding
a subject. So strong are the RBS neural circuits that even Jose Ortega y
Gasset was occasionally misled by them!
If this, then what
else? How can we expect
the uninformed masses to restrain their resentment and contempt for knowledgeable
people if even an academic is occasionally gripped by this primitive impulse?
In his The Revolt of the Masses (1930) Ortega y Gasset wrote that because
the 19th Century empowered the masses economically, the masses
were emboldened to view their undisciplined opinions, coming from within
themselves without the benefit of academic study,
to have equal validity compared to those from scholars. He writes: Today
... the average man has ... "ideas" on all that happens or ought to happen
in the universe. Hence, he has lost the use of his hearing. Why should he
listen if he has within him all that is necessary? There is no reason now
for listening, but rather for judging, pronouncing, deciding. There is no
question concerning public life in which he does
not intervene, blind and deaf as he is, imposing his ‘opinions.’ "
To the extent that
the newly empowered and unschooled mass of men influence public opinion (see
Price, 1970 and Allen, 1989), new forces are returning the pendulum "inward"
where today's fashion claims Truth can be found. Seeking truth by looking
inward is a dangerous trend. It contrasts with the scientific approach of
looking outward: observing, speculating, and testing the speculations by
appeal to additional observations designed to possibly falsify the speculation.
The scientific process
can lead to unexpected, unwanted insights. Thus, LBS inquiry represents a
threat to RBS beliefs, and naturally RBS people react by restricting LBS research.
Many subjects still cannot be studied because they are too threatening to
RBS beliefs. Some of the most lamentable example today are: 1) race differences
(Rushton, 1995), 2) the genetic basis of intelligence (Jensen, 1973), 3)
eugenics, 4) the sociobiological basis for rape (Thornhill and Palmer, 2000),
5) the role of sociobiological "group strategies" to account for Jewish history
(MacDonald, 1998), and 6) the origin of religious belief as an evolutionary
adaptation (Alper, 2000).
Scientific inquiry
into "uncomfortable" subjects is suppressed by RBS people, such as the (take
a deep breath) “Jewish, Marxist, politically-correct New York intellectuals”
who have already hijacked a few choice nuggets of our culture and who continue
to try to remold our values. New Age "feeling" people may be viewed as harmless,
but their politically correct intellectual comrades are harmful, for every
time they interfere with "thinking" people there is an incremental loss in
the prospects for civilization's continued growth and prosperity.
The Mutational Load
Theory
In most contemporary
primitive cultures women have an average of eight babies. Allowing for the
fact that in prehistoric times women may have
died during their reproductive years (due to difficult births, disease, starvation,
tribal warfare, etc), 50% to 75% of offspring would die before reaching reproductive
age (which was 16 or 17 years until modern diets changed it to its present
12 or 13 years). In modern cultures women bear an average of two babies,
and both of them are likely to live throughout their reproductive years.
If this sounds like progress, read on!
According to Alexey
S. Kondrashov (1988, p435) "in modern human populations detrimental
mutations are probably accumulating faster than they are being eliminated
by selection" due to the survival of a larger fraction of newborns than
in former times. He postulates that normally there is a steady‑state between
the processes of mutational degradation and preferential survival of babies
with few deleterious mutations.
Now, pause again,
and take a deep breath! Anyone reading this far will sense that they're entering
politically incorrect waters! To rationalize your desire to draw back, to
recoil away from the uncomfortable implications that you suspect are implied
by the preceding, you will probably feel obliged to question Kondrashov's
allegation: "Surely, deleterious mutations
do not occur at high enough incidence to matter in just one generation!"
But what if they do? Because if they do, and if humanity's slow march forward
toward an ever‑higher and more glorious pinnacle has depended upon the "sacrifice"
of those bearing the deleterious mutations, then what Pandora's box have
we opened by creating a society that assures survival for virtually everyone?
The implications
are unthinkable! "Would God permit a world that produces an ever‑expanding
population of people afflicted by physical deformities, physiological weaknesses,
and mental defects?" "If we are in dysgenic decline, is our only recourse
that reviled, discredited, mean‑spirited theory called eugenics?" "Didn't
Hitler practice eugenics?" (For the record, the answer is “no” – Jews score
higher on IQ tests than non-Jews.)
Theory predicts
that a population's genetic quality should decline when the mutational load
is not removed by a process of differential survival of individuals before
they reach reproductive age. This should be self‑evident (but I can provide
an analysis to support it, upon request). Is there evidence for a decline
in genetic quality during recent generations? The published literature has
little to say about this, possibly because it is a taboo subject. Epidemiologists
are still puzzling over the dramatic increase in asthma during the past few
decades (they say you can't blame it on air pollution). And the incidence
of "learning disabled" children is increasing rapidly (although this may
be due to changing thresholds for labeling children as such). Herrnstein (1978)
has studied IQ in America and concludes that
it has declined 4 or 5 points during the past 5 or 6 generations. However,
Herrnstein attributes this decline to a higher fertility rate for people
with low IQ, and this explanation is unrelated to mutational load theory.
Most traits are a product of both genetics and environment, and this presents
a special challenge to anyone seeking evidence for the case that we are in
dysgenic decline due to an accumulation of a load of genetic mutations.
As an aside, the
"Flynn Effect" (Flynn, 1987) purports to show an IQ increase of between 11
and 18 points between 1950 and 1980 among young Americans, with similar upward
trends in other countries. However, questions have been raised about the
use of the early, original IQ tests for measuring changes across generations,
and Flynn himself has concluded (1990) that this is the case. Interestingly,
IQ scores over time are rising while SAT scores and academic performance
are decreasing. These waters are muddied.
With the observations
of IQ trends ambiguous, we are left with mere theory to guide us. As there
seem to be no publications that address this question, I developed a very
simple model meant to explore how far a heritable trait can be "degraded"
when mutational load is not "corrected" by the normal amount of differential
survival of offspring. I used IQ, since it's a well‑studied, multi‑gene trait,
and it has a high heritability (0.75 according to identical twin studies
conducted by Bouchard et al, 1990). Allowance was made for
the fact that the average offspring IQ is a 70/30 weighting of the parent's
IQ and the genomic average IQ (defined to be 100) according to various studies
that start with Galton (1909). If all of the bottom 50 % of offspring fail
to reach reproductive age, the average adult IQ of the adult offspring would
be 105.7 in the absence of mutational load. A more realistic assumption is
to assume that survival probability ranges linearly from 0% at an IQ of
60, to 100% for an IQ of 140. This would produce an offspring average IQ
of 102.0. I interpret this to mean that the mutational load on IQ is about
2 IQ points! In other words, starting with a parent generation IQ that averages
100, the average IQ of offspring is 98, but after the 50% culling of preferentially
low IQ offspring during infancy and childhood, when the surviving offspring
reach adulthood their average IQ is restored to near 100!
Since this culling
effect is greatly reduced in developed societies, the theory predicts that
we are producing generations of offspring that reach adulthood with IQs that
average closer to 98 than 100. Over time this will lower the "genomic IQ"
(the IQ, which is commonly taken to be 100, toward which offspring IQ is
"attracted" with a 30/70 weighting). I calculate that this is a very slow
process.
If IQ is decreasing
by amounts as large as the Herrnstein study suggests, then it probably is
not due to mutational load. Herrnstein's theory of different fertility rates
for groups with differing "genomic IQ" may be capable of producing the faster
changes. That topic is not relevant to this essay, and will not be discussed
further here.
IQ is a multi‑gene
trait, and therefore has greater "inertia" to change from random mutations.
Single‑gene traits, such as hemophilia, should be capable of increasing their
representation in the gene pool at much faster rates. For example, before
modern medicine most individuals carrying the X chromosome allele for hemophilia
would have died before reaching reproductive age. Today, they can be treated,
as almost all of them are, and they then have the full potential of reproductive
years for passing on the hemophilic gene to their offspring (by way of a
sex‑specific X chromosome transmission path). Under such conditions the hemophiliac
gene can rise to levels in the genome that are much higher than had ever
existed in earlier times. (Although most cases of hemophilia can be attributed
to an inheritance of it based on family history, some cases are apparently
due to "spontaneous" mutations at a mutation-prone site.) This qualifies
hemophilia as an example of mutational load.
As more diseases
become identified with DNA locations, the list of examples of mutational
load genes will grow. The importance of addressing this problem will also
grow, not just because we will be able to identify more diseases as gene‑caused,
but because the incidence of gene‑caused diseases will be increasing and
the burden to society will become more costly.
The mutational load
theory, with its prediction of a deterioration of the mental and physical
health, or vigor, of the population, is just one more pressure on a civilization’s
continued existence. The timescale for repairing the damage that could be
done to the genome by a “too kind” society is much longer than the timescale
for creating the problem. Thus, the wealthy society unknowingly, and with
the best of intentions, contributes to its own decline. In other words, a
society that experiences a rise in wealth is hit by two new challenges: the
poorest in society are encouraged to become parasitic and the wealthy are
burdening society with defective children.
Empathy for the
plight of one’s children is a "micro-motive" produced by the genes. The "macro-behavior"
resulting from these micro-motives may produce a perpetually recurrent oscillation
of gene pool vigor, producing the rise and fall of civilizations. The oscillation
will not be sinusoidal, for the building process should take far longer
than the destructive process. How fortunate we are to be living through
the cusp of our civilizations apex! Yet how disheartening
it will be for our children who will live through the crash.
Why are the opinion shapers of society loathe to talk about
this? Every year that society postpones dealing with the problem is a year
that the problem grows, and becomes more difficult to solve. It's not just
mutational load; there are many other subjects deserving attention that are
not being addressed by responsible societal discourse. Some subjects are
just naturally taboo!
In this chapter
several factors were suggested as possible contributors to the decline of
a civilization. The next chapter provides some evidence that civilizations
rise and fall in the manner described in this chapter, and that today’s global
civilization might be the last one to exist because humanity is doomed to
be become extinct during the current millennium.
─────────────────────────────────
CHAPTER 19
─────────────────────────────────
FACTORS INFLUENCING THE FATE OF
CIVILIZATIONS
PART V: FASCISM
"Democracy
is the theory that the common people know what they want - and deserve to
get it good and hard!" H. L. Mencken
Any discussion of
causes for the collapse of an empire and threats to civilization would be
remiss if it did not include an analysis of what’s happening right now to
the American Empire. That is the subject for this chapter. The risk in describing
a current event is that it will inevitability lack
perspective, and it is fraught with the temptation to insert opinions instead
of facts. I have tried to prevent this chapter from becoming a “political
rant” but I know it still has that flavor. Forgive me for caring passionately
for my home country.
America went from being
the world’s biggest lender nation (peaking in 1981) to the biggest debtor
nation in less than 2 decades (passing from creditor to debtor in 1984).
When this happened, accompanied by a host of other troubling changes, everyone
should have been asking “why.” Interesting forces must have been at work
to account for such a switch. Can these forces be understood well enough to
assess the possibility that America’s economy and stature
as an empire are on a path to collapse?
The chapter reviews
American history with frequent reference to two theoretical sub-themes. My
hope is that the two theories will be supported so convincingly that when
my historical review arrives at the “present” I can use the theories to
extrapolate America’s future.
Theory 1: Review
of Group Selection Theory
This section will
review Chapter 11’s discussion of a sociobiological group selection theory
used to account for changes in individual behavior as conditions fluctuate
between war and peace. It serves as essential background for understanding
a trend in America that began in
the 1980s.
One enduring principle
of evolution is that the strong rarely miss an opportunity to exploit the
weak. The exceptions are few, but significant. The strong refrain from exploiting
the weak in three situations: 1) when related individuals are involved, 2)
among friends with a history of reciprocal relationships, and finally 3)
when one’s tribe is at war with a rival tribe this exploitation is rare for
intra-tribal interactions. This last situation was described in Chapter 11
in great detail, and it will be reviewed here because it is a crucial ingredient
for understanding America’s current predicament.
When a tribe is
at war with a neighboring tribe individuals are “programmed” to act altruistically
by becoming unquestioning patriotic warriors. Individuals also refrain from
any intra-tribal behaviors that are likely to weaken the tribe, and jeopardize
the outcome of the ongoing conflict. Any tribe in which this does not happen
cannot survive, so through a “group selection” way of thinking we expect
that all tribes that survive in such an environment will consist of individuals
that act this way.
After the tribal
conflict is resolved, perhaps with one tribe achieving decisive victory
over its rival, inter-tribal relations will be peaceful. The victorious
tribe may in fact achieve such overwhelming dominance over its newly enlarged
realm that it can be called an empire. During this prolonged interlude of
peace the need disappears for patriotic altruism. There is also diminished
need for intra-tribal good behavior since the overall strength of the tribe
will not affect the survival of its members. The intra-tribal component of
this theory can be summarized by stating that there are rewards for genes predisposing for intra-tribal niceness during warfare
and there are rewards for genes predisposing for intra-tribal
nastiness during peaceful conditions.
Since the alternations
between war and peace and war, etc., can often be shorter than a lifetime,
which is much too short for gene pools to evolve, human brains must be
pre-wired to produce these condition-based behaviors. This pattern for human
behavior is supported by game theory simulations (Choi and Bowles,
2007) and also a careful
re-reading of recorded history (Turchin, 2007). In order to understand
its relevance today I need to review how it came about at the transition
between human pre-history and recorded history.
Pre-History Lesson
Humans must have
lived in tribes for millions of years. Chimpanzees live in their version
of tribes, and they wage war with their neighbors, so it is not unreasonable
to assume that humans have lived this way since the time of the human/chimpanzee
split, about 6 million years ago.
The size of human
tribes was probably limited by a nomadic lifestyle as well as hunting and
gathering conditions, with numbers probably in the 20 to 50 range (chimpanzees
live in troops of about 10 members). Group selection pressures are probably
insignificant for such small tribes. Like chimpanzees, where males often
join neighboring troops, an individual’s fate is not determined as strongly
by the fate of one’s tribe when it is small.
Probably sometime
about 50,000 years ago, when tool making became a full-time occupation
for a designated artisan in most tribes, and when division of labor for
other tribal tasks may have begun, the size of human tribes must have grown.
Tribal cohesion would have greater rewards when this interdependence of
membership was important. A tribe could only sustain a large size when it
had specialized roles. And large tribal size was important when tribes
engaged in warfare. Whereas small tribes had been organized “democratically”
these larger tribes needed coordinated leadership to respond effectively
during inter-tribal warfare. I envision that tribal size was in the 50 to
150 size range from about 50,000 years ago until the end of the Pleistocene
(12,000 years ago). A tribal chief may have been identified for directing
warfare; during peaceful intervals the “chief” may not have had especially
important privileges, and decisions may have been made democratically.
During the Holocene
(starting ~12,000 years ago), which is an inter-glacial with a warmer climate,
there were new opportunities for supporting large tribes. Agriculture was
one path to growth, and marauding agricultural communities was another. Inter-tribal
warfare may have been more frequent and the fate of individuals may have
been tied more to the fate of one’s tribe. An intuitive understanding of
“all for one, and one for all” would have been an asset for all tribal members.
Greater powers would have to be given to the tribal chief, who may have
recruited his favorite and most loyal companions to help him during battle
and maybe during peaceful interludes. The democratic form of coordinating
tribal activities would yield to a more “leader controlled” tribal structure.
During the Holocene, as some tribes thrived and others were decimated, there
would emerge a new tribal structure in which a strong leader could count
on the loyalty and obedience of the others. Any tribe that embraced individual
rights and a deliberate process of reaching a consensus before taking any
action would simply disappear.
Theory 2: Emergence
of Strong Central Leadership During Holocene
Feudalism was a
form of social organization created to offer protection of workers (peasants)
by warriors who took their direction from powerful, land-controlling rulers
(kings) whose claim to power is based on their past performance as a warrior.
The king acted as if the land belonged to the kingdom that he ruled. He
allowed the peasant farmer to cultivate a portion of land in exchange for
a tax on the produce and sometimes an obligation to join with warriors when
called upon to defend the kingdom. This arrangement was made necessary by
the incessant threat of marauding bands.
Since it is “the
man with the hoe” where almost all production begins there is a sense of
unfairness inherent in feudalism. The warrior is not a producer, nor is the
king and his cronies. The peasant producers resemble a herd of cattle who are protected because they provide sustenance for
their keepers. It would probably be fair to say that the poor peasants are
exploited by those with power.
But for the powerless
peasant the deal may be good, even though there’s no alternative. By himself
he can’t protect his storage of crops, so a tax on them in exchange for security
seems fair. The land he tills can’t be protected from stronger neighbors
in search of greater production, so the arrangement of tilling an agreed
upon portion of the king’s land is a safe alternative. The underlying worry
that kept the peasant in his place was fear; the fear of marauders, the fear
of invasion by neighboring kingdoms and the fear of intimidating take-over
by his nearby fellow peasants. As long as this quasi-enslaved peasant works
hard he can count on surviving.
The willing peasant
is helped by having the slave’s mental attitude. Were he to rebel, as some
surely did, the alternative lifestyle would be risky and he would have fewer
grandchildren. Inexorably, over 12,000 years, a peasant class of men evolved
who instinctively accepted this feudal social order.
Feudalism has appeared
in many cultures, so the readiness to form a feudal society could almost
be called a “human universal.” Feudalism involves two mutually-dependent
populations: the masses of semi-enslaved, semi-impoverished peasants and
the minority of wealthy leaders and the defenders that they command. We should
not be surprised to find in 21st Century men a residue of readiness
for these two attitudes and a feudal form of governance.
American Revolution
The Founding Fathers
of America were fed up with England’s overbearing
rule. King George III tried to extract money from the American colonies.
When colonists objected to his excessive taxes he was enraged by their disobedience
and believed they should be dealt harshly. The “Tea Act of 1773” unfairly
favored the monopoly power of a British company, the East India Company.
The pattern of King George’s dealings with the colonists was clear, and there
was no hiding the fact that the colonists were being exploited in order to
favor the monarchy and a large British company. King George over-played
his hand and eventually lost everything at the conclusion of the American
Revolutionary War in 1783 (he was later called “the king who lost America”).
Thomas Jefferson,
the primary author of the Declaration of Independence (1776), wanted America to be an experiment
in democracy. He had no use for “allegiance to a monarchy.” Amazingly, some
colonists preferred to be ruled by King George III. They were referred to
as “royalists” and also “conservatives” since they wanted to “conserve” a
monarchy form of governance.
Jefferson and Alexander
Hamilton had a long-standing disagreement about the ability of ordinary people
to sustain a democratic form of government. Jefferson believed that educated
people could be trusted to govern themselves wisely, and that eventually the
world would rid itself of monarchies and dictators. Hamilton called this wishful
thinking because he believed human nature lacked the requisite wisdom to
sustain a form of democratic self-governance.
The Constitution
was written (1787) with safeguards against abuse of power. For example, it
included a ban on monopolies in commerce (prompted by their experience with
the East India Company). Many of its provisions were aimed at preventing
a repeat of the abuses suffered under the British monarchy. The creation of
three branches of government was supposed to achieve this. The Constitution
can be viewed as a reaction to a history of only monarchies and dictatorships
stretching back 2000 years to the first great experiment with democracy in
Greece.
19th
Century America
If history is viewed
as a swinging back and forth between power of the wealthy and power of
the people, then late 18th Century America was surely a swing
toward the latter. However, the late 19th Century had an opposite
swing. After the Civil War (1861-1865) some businesses grew powerful enough
to influence the US Congress, and undermine the moral attitude of the executive
branch. This time is referred to as the American “Gilded Age.” When it was in full swing (1877 to 1893) extravagant wealth
for the few contrasted with poverty for the many. The most memorable
wealthy few are referred to as “robber barons” (e.g., Rockefeller, Vanderbilt, Carnegie
and Morgan). They manipulated the federal government in ways that benefited
them greatly, with no regard for the workers who sustained their enterprises.
The government was too weak and unmotivated to prevent unethical financial
manipulations. Labor abuses led to uprisings and the beginning of unions.
This movement ushered in a “progressive era” (1896 to 1920). Slow progress
was made in the areas of worker’s rights (e.g., child labor
laws) and reversing other social injustice issues (e.g.,
women’s right to vote).
The Roaring Twenties
The 1920’s decade
has so many take-home lessons that are relevant for this chapter that I will
give it plenty of attention. As this section illustrates, history does repeat
itself!
The Progressive
Era is partly responsible for the booming economy of the 1920s. The Roaring
Twenties was a time of unleashed creativity, leading to many inventions,
new businesses, and advances in literature, art, music and entertainment.
America’s image of itself
is that this is where any entrepreneur could prosper and achieve the American
Dream. The optimistic belief that prosperity was in store for everyone overshadowed
the growing disparity of wealth between the rich and the middle class.
President Calvin
Coolidge (1923–1929) was naively optimistic about the economy as he proclaimed
“The business of America is business.” But
the so-called “Coolidge Prosperity” had not benefited everyone. The disparity
between rich and poor grew throughout the 1920s. By 1929 the top 0.1% of
Americans had a combined income equal to the bottom 42% (Brookings Institute,
1981). Businesses were not regulated sufficiently. Businesses, it seems, never
ask to be regulated. Speculation in the stock market was viewed by those
who had money to invest as the path to greater wealth. The stock market was
also not regulated sufficiently.
On October 24, 1929 the stock market
“crashed” and in 3 weeks had plunged 40%. During the ensuing Great Depression
the unemployment rate reached 25%.
According to Gusmorino
(1996) “The federal government contributed to the growing gap between the
rich and middle-class. Calvin Coolidge’s administration (and the conservative-controlled
government) favored business and as a result the wealthy who invested in
these businesses.” He goes on to write “…the Revenue
Act of 1926 … reduced federal income and inheritance taxes dramatically.
… In effect, [a wealthy person] was able to lower federal taxes such that
a man with a million dollar annual income had his federal taxes reduced from
$600,000 to $200,000.”
Sound familiar?
The Reagan to Cheney
administrations resemble the Coolidge administration.
There are other resemblances, and since this will play an important part
of this chapter’s argument I will use the rest of this section to paraphrase
key points in an article by Gusmorino (cited above, also at http://gusmorino.com/pag3/greatdepression/).
All quotes in the rest of this section are from Gusmarino.
Productivity increased
32% during the 1920s whereas wage earners only saw an income increase of
8%. “… the bulk benefit of the increased productivity
went into corporate profits…” “It was not that the surplus products of industrialized
society were not wanted, but rather that [the poor could not afford to
buy them, whereas] the wealthy were satiated [after] spending [only] a
small portion of their income.”
“Throughout such
a period of imbalance, the U.S. came to rely upon
two things for the economy to remain on an even keel: credit sales and luxury
spending and investment from the rich.” “One obvious solution to the problem
of the vast majority of the population not having enough money … was to let
[them] buy products on credit. The concept of buying now and paying later
caught on quickly. By the end of the 1920’s 60% of cars and 80% of radios
were bought on installment credit. … Installment credit … created artificial
demand for products which people could not ordinarily afford.” At the time
of the crash “80% of Americans had no savings at all.”
Sound familiar?
(The marketplace reinvented credit purchasing to maintain corporate sales
after President Ronald Reagan started busting unions, causing workers to
lose purchasing power – as described by Thom Hartmann on his radio show.)
“While America was prospering
in the 1920's, European nations were struggling to rebuild themselves after
the damage of war.” The US had loaned European
nations over $7 billion during the war, and during the 1920’s the rate of
loans rose to $1.25 billion per year. “Of these funds, more than 90% were
used by the European allies to purchase US goods.” One wonders if US corporations
were looking for markets to supplement the dwindling buying power of Americans
and cleverly influenced the government to subsidize them through a good-sounding
scheme of foreign aid.
After the crash
“the middle-class and poor stopped buying things with installment credit
for fear of losing their jobs and not being able to pay the interest. As a
result, industrial production fell…” (If Americans were smart they’d start
doing this now.)
Want to hear a
sanitized version of what caused the crash? Economists (whose infamous adherence
to the idiotic model that “individuals are guided in their personal decisions
by rational thought” should discredit most of what they say) tell us that
“The
Federal Reserve kept the money supply tight, because they feared the runaway
inflation that they saw devastating postwar Germany.
So they over-corrected. They should have let the money supply increase slowly.
And the other cause was the Hawley-Smoot tariff act, which was protectionist. The
word that the US was
about to tax foreign imports spread globally and caused other countries to
erect trade barriers, so everyone suffered from the loss of trade.” (thanks to Tom McDonough for relaying this.)
Before proceeding
with a recapitulation of the 1930’s recovery I want to highlight what I think
is the take-home lesson of what happened to the economy of the Roaring Twenties.
When corporations earned large profits they didn’t share them with workers.
This led to a shortage of people who could afford to buy those products.
The demand for products was there, but the ability to pay for them was not.
The marketplace was creative, however, for it promoted installment credit
purchasing. This stratagem had the temporary effect of maintaining purchases
and corporate profits. But optimism, based on years of a booming economy,
kept people from recognizing that the economy was really unstable and prone
to failure, until it actually failed.
No single company
can be blamed for the crash. Most of them behaved in their short-term interest,
oblivious to the possibility that they were “killing the goose that lays
the golden egg.” A better metaphor for the tragedy that’s possible when everyone
pursues their self interest would be Garrett Hardin’s “Tragedy of the Commons.”
In his 1968 article Hardin describes a hypothetical situation of a pasture
that is grazed by many flocks of sheep; each flock owner is faced with the
decision of whether to add more sheep to his flock. From the owner’s perspective
adding one sheep to a flock increases his income by an amount proportional
to his flock size increase. Since each owner thinks the same way they all
can be expected to increase their herd size. At some point the pasture’s
capacity is exceeded, and starts to become over-grazed. Still, it makes sense
for each flock owner to add one more sheep because the increment of overgrazing
from an additional sheep is small (being proportional to the total of sheep
grazing) whereas the benefit is proportional to the increment of the owner’s
flock size. Eventually the pasture is not suitable for any sheep, and all
sheep die. The lesson from this hypothetical exercise is that it is in the
long-term benefit for all flock owners to come together to create an agreement
concerning each person’s behavior so that all may continue to benefit from
the pasture’s potential. If we apply this lesson to the companies in the
1920’s they should have been regulated by a government that brought them together
to formulate a behavior that would promote financial survival of them all,
as well as any other social benefits – such as the welfare of American society.
In other words, market forces cannot be trusted to fix all problems, and
government is sometimes needed to provide solutions. Government is not “the
enemy” – it can be the solution. This lesson was either forgotten during the
Reagan administration (1981–1989), or it was intentionally ignored for the
short-term gain of corporate leaders with inexcusable disregard for societal
welfare.
There are so many
parallels to the present American economy that I had goose bumps while reading
parts of the Gusmarino article.
The underlying “human
universal” that accounts for this sad episode in American history is that
humans are capable of short-term greed; when the rich have the opportunity
to exploit the weak, they will.
The Great Depression
When Herbert Hoover
accepted the Republican party’s nomination for
President (1928) he declared “We in America today are nearer
to the final triumph over poverty than ever before in the history of any
land. The poorhouse is vanishing from among us.” In his inaugural address
March, 1929 (just 7 months before “the crash”) he continued this happy talk
with “Ours is a land rich in resources; stimulating in its glorious beauty;
filled with millions of happy homes; blessed with comfort and opportunity.”
It is likely that he believed his speeches. After all, “on the surface” the
economy was booming. But the waters were deep, and turmoil was rising inexorably
to the surface.
After the crash
Hoover reassured America that the economy
was sound. This is what leaders say when things falter. They’re not simply
trying to comfort their suffering followers, they probably actually believe
their optimistic pronouncements. After all, their beliefs are like a religion,
and it’s unthinkable that their religion could be wrong. Hoover’s religion was
Republicanism, or conservatism, which has at its core the idea that as
long as the wealthy aristocrats prosper the rest of the kingdom will be
OK. It’s what today is called “trickle down” economics. The mechanism by
which this trickle down is supposed to occur is called a “free market.”
Only someone with religious fervor could believe that the free market will
solve all problems. As Hoover repeated, we just
have to wait for market forces to right our economy.
Homelessness swelled
as bank failures wiped out savings and unpaid mortgages led to foreclosures.
Some people lived in old crates and shacks called shantytowns, which were
also referred to as “Hoovervilles” as a bitter reference to the fact that
Hoover refused to give
government assistance to the unemployed. He repeated his belief that the
economy was sound and that public confidence would soon lead to business
investments and job creation. But why should a business expand production
when their warehouse was full of merchandise that no one could afford to
buy? When this didn’t seem to work Hoover tried raising taxes
thinking that a balanced federal budget would help. But this strategy only
made things worse because it reduced discretionary income, as anyone should
have predicted. Hoover refused to believe
that it was right to help people outright because that would undermine their
self-reliance. Everything Hoover tried failed because
he only tried things that were compatible with his conservative economic
principles.
Again, we should
be reminded of the great Holocene experiment that favored men who embraced
a concentration of power at the top of the super-tribe. The king knows best,
especially during inter-super-tribe conflicts. During peaceful interludes
the notion of kingly infallibility was turned to a feudal economic order
for society, as if recovering strength for the next challenge from beyond
the kingdom’s borders. Human evolution is slow, and royal abuses of power
may not have been checked by mutations within the genome or the meme belief
systems. I like to view people as inclined to embrace one of the two major
approaches to how a society should govern itself. The original idea is that
everyone should be involved in reaching a consensus (e.g., the Japanese management
style). The other is that decisions should come from above, from leaders,
from the gods! Little people should not question the gods, nor should they
question the king.
I have no doubt
about my inclination! The king’s infallibility is as much
an illusion as all the gods that humans have invented. Both repositories
of wisdom are “imposters” – dangerous, malicious imposters. I also have little
faith in the consensus form of governance, for the same reasons as Alexander
Hamilton. Most humans lack critical thinking skills; they latch onto whatever
beliefs they are guided to by genetic predisposition and tribal taboos, and
they behave in ways that are a confusion of what was adaptive in the ancestral
environment and what seems to make sense within a modern paradigm.
Hoover’s inclinations
are obvious; he was so Republicanized that during his term as president the
market forces he believed in, the ones that he interpreted as signifying
a fundamentally “sound economy,” just kept making things worse. Most of the
world was also in its own economic depression, thanks to World War I and
minimal real help from America. Consequently,
in America’s time of need
there was no one to help. (I’ll come back to this point in a later section.)
Hoover was “king” and
he had no answers. The deepest depression year was 1933, when Hoover left office and
Franklin D. Roosevelt (FDR) took over as president.
It is ironic that
FDR was born into aristocratic wealth. His outlook should have been like
Hoover’s. But maybe he
had the pre-Holocene genes that distrusted concentrated power. Or maybe his
personal experience with “polio” changed his outlook, and sensitized him
to the suffering of others.
FDR showed that
government, when properly used, could be “the solution” just as starkly as
the previous administrations showed that government could be “the problem.”
If government didn’t try new things then unchecked market forces would simply
continue grinding down the little guy and a continuing depression would bring
the American experiment in democracy to an end. The common theme of FDR’s
“New Deal” was a shift from serving big business to serving working-class
people.
FDR moved quickly
to impose overdue regulations on banks and the stock market. Minimum wage
and maximum work hour laws were passed (June, 1933). Jobs were created by
the government under the Public Works Administration (WPA). The Social Security
Administration was created in 1935 to provide a minimum of financial assistance
during retirement, unemployment or disability. Labor unions were treated
with less hostility than during the Republican administrations. Other public
works were created, such as the Tennessee Valley Authority, which not only
created jobs they also improved living standards in the rural areas surrounding
the TVA. Banking was made safer by his creation of the FDIC. Throughout all
these massive government initiatives FDR was concerned about federal budget
deficits. This concern made him somewhat cautious, but a growing resentment
by Republicans put the brakes on his innovative programs starting in his second
term (1937).
There is still argument
whether FDR’s progressive programs were responsible for bringing America out of depression,
or whether the immense budget deficit spending and job creation demanded
by World War II was mostly responsible. However, there is no argument that
from 1933 onward America’s depression was
less painful than it would have been without FDR.
FDR provided strong
leadership to a nation in economic crisis that might otherwise have floundered
and faded from world relevance amid starvation and misery. His hopeful, moderate
policies also prevented people from flocking to the American fascists and
communists, who could have created a dictatorship. Where Hoover was a weak leader
who was clueless about what to do, FDR understood the essential nature of
what had to be done and he was persuasive in getting on with needed changes.
Part of FDR’s persuasiveness was his ability to appeal directly to working
people, to allay their fears, nurture hope and renew national optimism. This
is called “leadership.” The entrenched business community hated FDR, and
resisted his programs as strongly as they could. Even some of FDR’s friends
from the aristocratic wealthy class called him a traitor. FDR achieved success
by switching government’s main mission from partnering with business to partnering
with people. And under FDR “We the people” usually prevailed over wealthy
conservatives who resisted people policies.
Theory and Reality
During the 1920’s and 1930’s
Let’s pause for
a moment and consider if the Roaring Twenties and the Great Depression are
compatible with the two theories that are an underlying theme for this chapter.
The 1920’s were
a time of peace following a world war. According to Theory #1 patriotic altruism
during the war years should have been replaced by the pursuit of individual
fulfillment during the following peaceful years. A frequent theme of 1920’s
social commentary is that individual liberation and the pursuit of pleasure
became tiresome; a purposeless life emptied the soul of meaning. Ennui and
aimlessness, amid hedonistic diversions, pervade the literature (e.g., The Great Gatsby). The new rich felt no social obligation for
others. This, indeed, is what Theory #1 predicts should have happened during
the 1920’s.
The 1930’s somewhat
resemble a time of war, when most people are suffering as if besieged by
an outside enemy. Compassion is rediscovered; those for whom food and housing
are scarce receive help from those better off (but only from the residual
of the middle class, for prospering wealthy people were inclined to ignore
other people’s suffering). FDR tapped into a groundswell of dissatisfaction
with the laissez-faire policies of the previous decade when
he appealed in 1933 for people to vote for a leader who will ask for sacrifice
from those who are “able to help” lend a hand to those who are unable to
help themselves through no fault of their own. This altruistic response to
a national need is what Theory #1 predicts.
Theory #2 also fares
well when comparing these two decades. During the 1920’s regulations were
lax and the strong took advantage of opportunities to exploit the weak. Business
influenced government, creating a partnership that was so obviously corrupt
that muckraking journalism had a “field day” exposing it (e.g.,
H. L. Mencken). If corporations could have installed a figurehead king to
rule over America they would have
done it. The wealthy aristocrats would have welcomed an American counterpart
to King George III in order to more efficiently enslave workers and dominate
world commerce.
During the 1930’s
FDR might be likened to a king, considering his expansive use of executive
power to make bold changes. FDR might even be described as a beneficent dictator.
However, FDR respected the powers of the other two branches of government,
which did at times deal him defeat. More importantly, FDR worked to move
wealth and power from the wealthy few to the many in poverty. Since his actions
had the sanction of most Americans, it is fairer to say that FDR returned
the country closer to the pre-Holocene tribal democracy in which decisions
are made by consensus.
The 1940’s
The American involvement
in World War II technically began when Japan bombed Pearl Harbor (1941). Years before
this event FDR was concerned about the looming war in both Europe and the Pacific
but he was unable to mobilize the American people to prepare for it. Massive
war spending that began in 1942 gave a boost to employment and the economy
generally. Factories that had been building cars at a slow pace swiftly
switched to producing tanks for which there was unlimited demand. (With “full
employment” the WPA and other employment programs were discontinued.) War
material was shipped to the British (under the Lend-Lease Act) before America was officially
at war with Germany. FDR did not require
payment for these shipments, which contrasted with the way Coolidge pretended
to help European countries during the 1920’s. FDR died April, 1945, a few
months before the Allies won the war.
The Democratic Party
dominated American politics from 1933 until about 1968, thanks to the effectiveness
of FDR’s three presidential terms of office during the first 12 years of
this period. The middle class prospered throughout the 1940’s, 1950’s, 1960’s
and 1970’s, and FDR can take credit for that too.
The 1950’s
I have first-hand
knowledge of America starting in the
1950’s, for I started college in 1957. My family was middle class and I recall
the excitement of getting a television, a new family car every few years,
and a general sense that society was stable. The Cold War was always in
the background of our thinking, and the ever-present concern about atomic
warfare with Russia instilled in everyone
a serious side. Sputnik prompted the University of Michigan to offer an Honors
Program to help catch up with the Russians in space technology, which I
took advantage of.
Some of my college
reading assignments accused my generation of being disengaged from social
issues and political activism, and being overly concerned with individual
pursuit of material wealth. We were accused of being too willing to wear
a “gray flannel suit” and work obediently for a large company. In retrospect
this was merely the quiet before the storm, which was unleashed in the next
decade.
The Sixties
“The Sixties” is
sometimes defined as 1963 to 1973. I’ll treat the beginning as 1960.
General Dwight Eisenhower
was courted by both the Democratic and Republican parties before he decided
to run as a Republican (1952). The Eisenhower administration (1953-1961) has
stood the test of time. He was experienced in assessing intelligence reports,
and he correctly assessed Soviet missile capability as less of a threat than
popularly believed. He launched the Space Race with Russia by establishing
NASA (1958). He also initiated the building of a national freeway system
(in case the military were to need it).
Eisenhower was not
the traditional Republican, since he embraced much of FDR’s New Deal. He
expanded Social Security coverage and established the Department of Health,
Education and Welfare. He enforced school integration, prevailing over the
resistance of the Arkansas governor (1957).
In the 1950s the
president of General Motors captured the essence of how to promote corporate
interests when he said ”What’s good for General
Motors is good for America.” Perhaps this
famous quotation influenced Eisenhower as he wrote his 1961 televised speech
about the military/industrial complex. He feared that large industries that
serve military hardware needs would have so much influence over the government
that they would dictate spending decisions without being held accountable
to voters. By this time American corporations had been growing for two decades,
and they might have been emboldened to explore the limits of their growth
potential. One of those limits for factories is the cost of labor. At this
time workers were feeling pretty good about their prosperity, thanks to laws
that protected unions. A collision was unavoidable, and the tumultuous Sixties
were when it occurred.
If Eisenhower were
alive today he would expand his military/industrial complex fear to include
the influence of all large corporations, not just the military ones. Large
corporations share the same “personality” by making opportunities that benefit
them at the expense of anyone else sucker enough to subsidize them. The new
industries to fear include pharmacy, health, insurance, banking (including
credit), mortgage lenders – and others (discussed below).
The presidency of
Jack Kennedy (1961-1963) symbolized the rising influence of a younger generation,
a passing of the torch of power from an older generation (Eisenhower) to
a younger one. Eisenhower would always be General Eisenhower, the man who
led the Allies to victory in World War II, while Kennedy was all about future
possibilities. Lyndon Johnson’s presidency (1963-1969), which began after
Kennedy’s assassination, succeeded in passing many of Kennedy’s progressive
policies.
I recall remarking
to a friend in the mid-1960’s that this time would be remembered as the best
of all times in both American and world history. America was prosperous,
the Cold War had not produced a nuclear holocaust, and there were indications
that in the future American society might coarsen and become unstable. I
believe that subsequent history has confirmed my assessment.
Prosperity in America just kept growing
after World War II. Babies born after the war were raised by parents who
remembered the Great Depression and wartime sacrifices, and they vowed to
save their children from these deprivations. There was a temptation to “spoil”
the children, and those born between 1945 and 1950 were college age starting
in 1963. It is more than a coincidence that this is when college age students
exhibited an excess of irresponsibility, a mocking of their parent’s work
ethic and a rebellion against conformity. Drugs, hanging out, mocking society
and protesting anything embraced by the Establishment created a generation
gap like no other.
Gone were the accusations
hurled at my generation, those born just before the war and during, that
we were too quiet and disengaged from public affairs, or that we were preoccupied
with material wealth, or a desire to work obediently as a Company Man. I
was fascinated by the undisciplined, free-thinking lifestyle of those just
a few years younger than me. The questioning of authority, and especially
the government, became a constant irritant for the Establishment, comprised
of a generation that came immediately before mine.
Being in that transition
generation, between those older than me in the Establishment and the younger
hippies, predisposed me to see both sides of the arguments. I continued my
work on the space program with NASA government “establishment” funds while
socializing with some of the hippies. My sympathies were divided, for although
I could not endorse irresponsible lifestyles I nevertheless agreed with the
younger generation about one of the most important things that irritated
them: a compulsory draft that forced peace-loving young men to take-up arms
for killing foreigners who
didn’t threaten America. The entire Vietnam
War struck me as immoral. The so-called “domino theory” neglected nationalism
considerations, borne out by subsequent events, so one alternative explanation
that I found attractive was that the war benefited the military-industrial
complex.
It is probably always
true that whichever class of a society is favored by the greatest growth
of wealth, that class will increase its influence in changing the society.
During the Sixties, as before, the middle class grew in wealth and influence,
and it is natural that this would scare the Establishment. Other things besides
losing influence must have scared the Establishment; the very stability of
society appeared to be threatened. Just consider that political assassinations
felled Jack Kennedy, Martin Luther King and Bobby Kennedy. The disenfranchised
of society rioted in Los Angeles, Newark and Detroit. Whereas during
the Great Depression there was cause for concern over the country’s survival
due to an economy that seemed resistant to cure, during the late Sixties
there must have been the same concern that was instead based on growing social
instability.
This concern over
social instability may have been a factor in the election of the first true
Republican since Herbert Hoover, Richard Nixon. He was elected president
in 1968, the year of the aforementioned race riots and assassinations. Nixon
promised to end the war in Vietnam and achieve social
stability by restoring “law and order.” Barry Goldwater had sparked a resurgence
of interest in reviving Republican control of government 4 years earlier,
but when he campaigned (1964) the hippy/Establishment culture war had only
begun and was not a paramount issue for voters. Goldwater rejected the New
Deal progressivism that had brought America out of the Great
Depression. Although Nixon won the election largely over a desire to end
social instability, he was also a beneficiary of what Goldwater started.
The Seventies
Nixon was accused
of being an Imperial President (Schlesinger, 1973) because of his exercise
of more power than allowed by the Constitution. His original transgressions
were intended to bring the Vietnam War to an end by expanding the war to
nearby countries without the required Congressional knowledge or approval.
Later Nixon expanded his powers to domestic issues. Congress passes budgets
and the president can sign or veto them. Nixon assumed powers not in the Constitution
to reward and punish those who agreed and disagreed with him. He also selectively
enforced laws. Some of the Nixon administration offences include “burglary,
forgery, illegal wiretapping and electronic surveillance, perjury, obstruction
of justice, destruction of evidence, tapering with witnesses, giving and
taking bribes and conspiracy to involve government agencies in illegal actions”
(wikipedia). His inept cronies were caught illegally wiretapping Democrats
at the Watergate Hotel, and this led to an unraveling of his presidency and
his resignation (1974).
Many aspects of
Nixon’s presidency resembled rule by royalty, by King Richard Nixon. The
Constitution was written to prevent what Nixon tried to get away with; thank
goodness for the ineptness of his cronies!
Gerald Ford was
Vice-President when Nixon resigned, so he completed Nixon’s term as president
(1974-1977).
Probably because
Nixon had “blown it” for the Republicans the election of 1978 went to Democrat
Jimmy Carter. I believe his presidency (1977-1981) has been unfairly maligned,
and I suspect this is due to a re-writing of history by Republicans. Carter
is the only US president to win
a Nobel Peace Prize (2002). He created a Department of Energy, and promoted
conservation and new energy technology development for the goal of achieving
energy independence. During his administration oil imports were reduced
50%. He is also known for his human rights activism.
Carter’s interest
in achieving oil independence was largely a response to the 1973 OPEC decision
to reduce oil exports in order to increase profits from the ensuing shortage.
This occurred during the Nixon administration, but Nixon never addressed
the matter. Peak oil production in America occurred in the
middle of the 1970’s, so American oil producers were handicapped in ramping
up production to stave off rising oil prices. The economies of 20th
Century advanced countries ran on energy produced by oil, which mostly
explains why Carter’s administration was burdened by high inflation, high
unemployment, slow growth and an increasing federal budget deficit. A “malaise”
pervaded the country near the end of Carter’s term. In short, OPEC ruined
Carter’s presidency.
The Reagan Era,
the 1980’s
President Ronald
Reagan is the protagonist of this chapter. He aspired to be the Coolidge
of the 1980’s. His attempt to unflinchingly pursue conservative policies that
threaten to destroy the middle class ran into just one problem: the rest
of the world was prospering and had money to loan credit companies so that
Americans could buy, buy, buy until they dropped – in 2008.
Well, I’m getting
ahead of my story, so let’s start over with Reagan’s election in 1980. Reagan
won the election because Carter was pursuing long-term solutions and Americans
demanded short-term solutions. When Reagan said “It’s Morning in America” that’s just what
voters wanted to hear as a counter to the Carter malaise. Reagan was charming,
he could speak well, he made jokes, and never
mind that he set in motion ruinous policies as Governor of California.
As governor of California he was cheered
by voters who liked his pledge “to send the welfare bums back to work” and
“to clean up the mess at Berkeley.” In order to lower
taxes he disbanded mental hospitals, which had the effect of filling the
jails with people the jail staff weren’t prepared to deal with.
Reagan’s presidential
campaign echoed his California governorship, whose
disastrous consequences were not apparent to national voters. He stated that
“Government is not the solution to our problems; government is the problem.”
What a clever line, for anyone who can’t think. He therefore won the election
(1980).
Reagan was a conservative’s dream-come-true. Half a year into his administration
the federal air traffic controllers went on strike, so he fired them, thus
sending a message to employers that they could confront unions. Also during
his first year in office he lowered taxes by amounts that ranged from 20%
for the wealthiest to 3% for the lowest income bracket. The underlying theory
for this imbalance is called “trickle-down” economics, which assumes the
wealthy will invest their savings in job-creating businesses which will
benefit the poor. An alternative explanation for tax cuts that favored the
wealthy is to consider who funded his campaign then recall the old saying
that “you dance with the one that brung ya.”
In 1986 Reagan signed
a second tax reducing bill, which lowered the top tax rate from 50% to 28%
while the bottom rate was raised from 11% to 15%. In addition, interest on
credit card debt could no longer be declared as a deduction, which hit poor
people harder. Capital gains were taxed at a lower rate, which helped wealthy
investors. The same tax act increased incentives for investing in home ownership
versus home rental, which also favored the wealthy. Changes were made to
the Alternative Minimum Tax that had the effect of increasing taxes for the
middle-class, with ever-increasing effect as inflation raised incomes.
What an amazing
feat for Reagan to have lowered wealthy taxes from 70% to 28%, while raising
rates at the low end of the income scale, and all in just 5 years! Did this
Reverse Robin Hood tactic improve the American economy? While it’s true that
the rate of inflation and jobless rate came down, tax revenues also came
down at the same time that federal expenditures went up. The budget deficit
just kept getting worse during the Reagan years. The federal government had
to borrow to pay for the national debt, which grew from $0.7 trillion to
$3.0 trillion during his presidency.
A hallmark of his
administration is that he did what corporations wanted: union-busting,
less government regulation of business, lower taxes for the wealthy and
disregard for anti-trust laws. Working people started losing their good-paying
jobs and had to settle for lower-paying ones, median income stagnated and
eventually decreased, employee benefits began to erode (health insurance
and retirement funding). Families headed by two wage-earners became the
norm, and job security worsened. Downsizing while promoting
overtime allowed employers to save on health care costs. The working
classes became docile for they feared that complaining could jeopardize
the jobs they had. Social stability was achieved, and labor costs for corporations
were reduced. Corporations achieved some of these savings by lobbying lawmakers
and infiltrating the executive branch of government. Corporate mergers were
allowed to happen in defiance of monopoly laws. “Trickle down” didn’t happen,
because corporations aren’t motivated to help workers, they’re motivated
to exploit them, wherever that exploitation is easiest and cheapest. Instead of “trickle down” the American economy had become
“trickle up” – with the wealthy able to steal from the poor.
In 1981 the US was at its peak
as a creditor nation. During the last six Reagan years America became the world's
largest debtor nation, with an economy increasingly owned and controlled
by foreign investors from across two oceans. In the 23 years since 1981 America changed from having
a $3.3 billion credit standing to having a $2.5
trillion debt owed to mostly foreign investors. The turning point from creditor
nation to debtor nation occurred in 1984, just 3 years after Reagan took
office. (Reader, judge for yourself whether President
Reagan deserves the nostalgic accolades that Republicans heap upon him as
they campaign against Democrats in 2008.)
One of the most
interesting economic tricks of the 1980’s relates to consumer credit card
purchasing. As Thom Hartmann explains (on his radio shows), the immediate
effect of busting unions and creating opportunities for businesses to exploit
ways to lower wages is that ordinary consumers had less money for buying
the products businesses were trying to sell. This situation resembled the
Coolidge years, for the same reasons, and some clever people recognized this
as a business opportunity. Why not restore credit purchasing by offering credit
cards (the new technology twist) for “buy now, pay later.” It worked during
the 1920’s, so let’s try it again. The credit industry grew, and grew, until
today it is a $30 billion dollar industry. Personal savings during the past
25 years have undergone the same changes that occurred during the 1920’s,
from positive to negative. Investors from around the world have come to America to keep the credit
industry funded, thinking that this was a good investment opportunity.
And it has been a good investment opportunity, for the government has allowed
the industry to charge “usury” rates.
I stated earlier
that Reagan aspired to be the Coolidge of the 1980’s. You may wonder why
Reagan did not produce an economic collapse like the Great Depression. The
answer is that during the 1920’s the rest of the world was still suffering
from World War I and they could not loan money to alleviate the American debt
and domestic credit crisis. During the 1980’s the rest of the world was prospering,
and they could help Reagan continue his fiscally irresponsible policies,
and they could buy American credit card debt. This postponed the day of reckoning
by 25 years, as I will describe in a later section.
There’s another
way of viewing Reagan if you are most interested in a long-term campaign
of returning America to that conservative
dream of being ruled by a wealthy elite. The shortest path to “royal rulership”
is to ally the government with corporations and create policies that benefit
corporations. There’s a small matter of disregarding common people. The
trick in doing this is to fool the middle class into thinking the government
is doing things for them. Instead of declaring honestly that measures are
being taken to “starve the beast” that helps the needy by creating a debt
that requires drastic reductions in social benefits, the conservatives pitch
the merits of individual ownership and getting the government out of the
way for economic growth. As corporate lobbyists draft laws that benefit their
corporate employer, and contribute to the campaigns of congressmen who pass
those laws, the government is transformed from being “of, by and for the
people” to one that is “of, by and for corporations.” There’s a name for this
kind of government; it’s called fascism!
Benito Mussolini
defined fascism as “…a system of government
that exercises a dictatorship of the extreme right, typically through the
merging of state and business leadership, together with belligerent nationalism.” I’ll discuss his
references to “dictatorship” and “belligerent nationalism” in a later section
(“The Cheney Administration”). For now, focus attention on the reference
to “merging of state and business leadership.” In the shortest path to royal
rulership the first step is to form an alliance with corporations, and this
was Reagan’s greatest accomplishment.
I hereby confess
to voting for Republicans until I saw Reagan’s actions as president during
his first term. For his second term election I started voting for Democrats.
This year, 2008, I will not vote, for reasons given in the last section.
The 1990’s
I have noted that
Democrats dominated American politics from FDR’s election in 1932 until Nixon’s
election in 1968. Since then American politics has been dominated by Republicans.
Reagan was wildly popular during the election following his two terms, in
1988, when George H. W. Bush (Bush Senior) won. Bush had been Reagan’s vice-president
for 8 years and the American people were clueless about the slow-ticking
economic time-bomb that this administration had created.
Bush Senior had
a single term presidency (1989-1993). His first task was to get control of
the national debt, that had tripled during Reagan’s
tenure. He and the Republicans wanted to reduce spending while Democrat-controlled
Congress wanted to raise taxes. His efforts to do both ended with more tax
increases than budget cuts, and this caused Republicans to feel betrayed.
This set the stage for Bill Clinton’s election win in 1992.
The Bill Clinton
Years
Bill Clinton’s presidency
(1993-2001) began just as the American economy was recovering from a mild
recession that kept Bush Senior from being re-elected. During his first year
in office he supported and signed NAFTA (North American Free Trade Agreement).
This may be one of his gravest mistakes, for it accelerated American job
losses. His administration wanted to reform health care but well-organized
lobbyists (AMA and health industry) got in the way. One of his successes
was lowering taxes for the poor and raising them slightly for the wealthy.
During most of his two terms the Republicans controlled Congress, and this
hampered what he could accomplish. Nevertheless, the
economy grew dramatically during his second term due largely to the internet
and speculation in the high technology sector. By the end of Clinton’s presidency the
federal budget was balanced and the national debt had been paid off, leaving
a surplus. Al Gore was vice-president for both terms and he played a major
role in reducing the size of government. It’s ironic that the Democrats were
able to accomplish two key goals that Republicans claim as theirs: reducing
the size of government and balancing the budget to the point of creating
a surplus.
The Cheney Era
The election of
2000 was nasty by anyone’s account. Karl Rove had refined the winning use
of dirty and divisive politics from the days he helped George Bush Junior
win the Texas governorship. Rove
had set as a goal delivering Texas to the Republican
party, and he was amazingly successful in this.
His tactics were so unethical that even George Bush Senior kept him at arm’s
length. The Rove-orchestrated smear of Senator John McCain during the Republican
primary campaign was adept, and it knocked McCain out of the running. Even
ignoring the voting irregularities in Ohio and Florida, it is a credit
to Rove that he propelled Bush to a close-call
vote against a more articulate, more experienced and more intelligent Al
Gore. The outcome was eventually decided by the U.S. Supreme Court.
Because subsequent
events have shown that vice-president Dick Cheney
has played a very important role “behind the scenes,” and some think his
influence exceeded Bush Junior’s, I will refer to the following 8 years as
the Cheney Era. Bush Jr. acted as a figurehead, or, as the campaign touted,
someone “regular folk” would like to have a beer with.
The Cheney Era has
been the least concealed attempt to recapture “royal rulership” of any previous
Republican administration, including the one that created the Great Depression
by Calvin Coolidge. It is generally recognized that the Bush family has a
history of elitism (Philips, 2002), so it was not surprising when Cheney began
maneuvering to expand the power of the Executive branch of government. Cheney
uses the term “unitary executive theory” to justify giving the president final
decision over all actions by the Executive branch of government. Other behaviors
of the Cheney administration expand on this, the most egregious of which
are “signing statements” that specify what parts of a newly passed law by
Congress the president intends to execute. Every year of their two terms
has seen a growth of presidential power under Cheney.
The federal budget
surplus that the Cheney administration inherited was returned to the taxpayers
in the form of a $1.35 trillion tax cut, which almost exclusively benefited
the wealthy – especially when secondary effects are taken into account. Cheney
sold the tax cut with the following slight of words “the surplus is not
the government’s money.” Because a recession was looming Treasury Secretary
Paul O’Neill opposed the tax cut, claiming it would worsen the budget deficit
and threaten Social Security. But if that was one of the purposes for the
tax cut, O’Neill’s argument must have just strengthened their resolve to
go through with them – which they did. O’Neill describes a conversation with
Cheney thus: Cheney says to O'Neill, "Reagan proved that deficits
don't matter. We won the midterm elections. Our due is another big tax cut.
(from the 2004 book The Price
of Loyalty by Ron Suskind, about Paul O’Neil’s rocky tenure as Treasury
Secretary). The federal debt rose every year of the Cheney administration,
to a current value of $9.5 trillion.
Cheney wanted to
loosen regulation of industry, and two of those to benefit were the mortgage
and investment industries. Sub-prime interest mortgages were promoted during
the second term (Federal Reserve Chairman Alan Greenspan did not object)
and they were issued to home buyers regardless of ability to pay (some without
jobs or down payment), and the mortgage loans were bundled and sold as highly
rated “derivatives.” When owners began to default on mortgage payments and
their property was foreclosed (starting in 2007) there was confusion about
who owned the property. Executives of mortgage companies guilty of unethical
financial dealings nevertheless collected large salaries and bonuses, even
as some of those companies went bankrupt.
Much of the Cheney
“royalization” project was done using lobbyists who drafted legislation that
Congressmen would promote in exchange for campaign contributions. There are
many examples; consider the following.
The Medicare Act
of 2003 created a drug insurance program to be offered by private insurance
companies that would compete for the privilege of offering this supplement
to Medicare. It passed in Congress after a long, arm-twisting session with
irregular procedures, well past the midnight hour, with a provision
that the price of drugs could not be negotiated by the private insurance
companies. This law, which has greatly benefited the pharmacy industry,
is the kind of legislation that the pharmacy lobbyists would have drafted
if they could have, and they most likely did.
Cheney must hate
Social Security. He began with a first step for its dismantlement the idea
of allowing contributors to divert part of their Social Security payment
obligation to a personal account in the stock market. This was given the nice-sounding
name of “personal ownership.” But since it was promoted at a time when the
stock market was faltering the idea aroused broad public disdain and it went
nowhere. This legislation would have greatly benefited investors on Wall
Street, and it might have been drafted by their lobbyists.
Early in the first
term Cheney convened a secret Energy Task Force meeting to create an energy
policy. The list of participants has never been released, but given its supply-side,
oil oriented structure one can speculate that oil company executives were
in attendance.
The Cheney position
on global warming, which they slyly refer to as “climate change” (thanks
to Frank Lunts), is that it may be a natural climate cycle so it is premature
to enter into any international treaties (e.g., the Kyoto Protocol) that
would restrict CO2 emissions or regulate industry.
Words can deceive,
and that’s what was done with the “Clear Sky Initiative” (another sly title
that misleads), an amendment to the Clean Air Act (that Cheney’s administration
reluctantly inherited). This regulation-emasculating legislation failed to
be enacted by Congress so Cheney implemented key provisions by instructing
the EPA (Environmental Protection Agency) to proceed with NOx, SO2 and mercury
trading; this questionable administrative procedure led to a court challenge.
The EPA systematically
censored reports it was mandated by Congress to issue in a way that made
it seem like industry regulation was not necessary. Climate scientist Jim
Hansen was ordered to not speak candidly at public scientific meetings because
his message was clearly at variance with what the Cheney administration wanted
to believe. Even NASA is guilty of implementing some of the administration’s
“clearance before speaking” directives. It is easy to imagine that the oil,
coal and electric utility industries lobbied for these policies.
Every business must
like cheap labor. There’s nothing wrong with this, but is it legal to urge
the Executive to hamper enforcement of illegal immigration in order to have
a large pool of compliant workers? The “undocumented alien” (UDA) will not
complain about job safety, overwork, low pay and
they will not unionize. The Cheney attitude of neglecting to enforce employer
sanctions was a “dream come true” for the poultry, meat packing, agriculture,
construction and landscaping (gardening) industries. The Democrats in Congress
also like illegal immigration because Mexicans tend to vote Democratic. The
assumption is that after hordes of Mexicans make it to America they will become
American citizens through amnesty legislation, which most Democrats and
Republicans want. But the UDAs create a burden on local communities, for
they use emergency rooms for their routine medical care, they have lots of
children who attend schools paid for with property taxes, they steal people’s
identification (mostly Social Security numbers), they cause high-speed chases
that lead to frequent deadly accidents and they trash rural pathways to their
transportation rendezvous sites. I live 7 miles from the Arizona/Mexico border
and I understand these problems. The Republican business people are as guilty
as the Democratic Party for neglecting these problems.
Within days after
the 9/11 Saudi Arabian Islamic terrorists flew airplanes into the World Trade Center buildings and the
Pentagon, Senator John McCain renewed his call
for attacking Iraq. He advocated bombing,
not an invasion at that time, and his rationale was to scare Middle Eastern
countries into “not messing with America.” He didn’t claim
that the terrorists had been helped by Iraq’s Saddam Hussein.
Cheney, Rumsfeld and Bush, however, had been talking about invading Iraq since the first
month of the Cheney administration, and their plan was for an invasion. The
real motivation for such an invasion may never be made public, but speculation
includes a desire to have access to Iraq’s oil fields (a
boon for oil company business). The 9/11 attacks gave Cheney and Rumsfeld
the excuse for invading that they wanted. Cheney heavy-handedly interfered
with the CIA investigation of any possible links between the 9/11 attackers
and Iraq, and made unprecedented
visits to the CIA that are described by intelligence officials as intimidating.
Selling the war took 1½ years, and the final step was to convince the United
Nations that Iraq had weapons of
mass destruction poised for use in ways that would interrupt Saudi Arabian
oil production and lead to global recession. Cheney’s aide “Scooter” Libby
drafted a speech for Secretary of State Colin Powell to deliver at the UN,
with only a few days left for fact-checking. Powell’s credibility was exploited
shamelessly, and he must view his speech before the UN as the most embarrassing
day of his life. The speech did the job, for the UN passed a resolution,
which was Cheney’s “green light” for the invasion.
The FDA (Federal
Drug Administration) is supposed to regulate the pharmaceutical industry.
Instead, they protect and promote it. Drug testing, that used to be conducted
by independent laboratories, is now conducted by the pharmaceutical company
that manufactures the drug. As a result drug side effects are usually discovered
after it has been on the market and billions of dollars have been made from
it. Cheap alternatives to drugs are disparaged by bogus experimental trials.
When a patent drug approaches the date when it should become “generic” the
FDA allows a small change to be made
that resets the calendar. It has been estimated that over
100,000 deaths can be attributed to drugs that are taken as prescribed by
a doctor (to the extent that charlatans are wearing white coats the above
statistic may be more of an indictment of doctors than the pharmaceutical
industry.)
There are so many
regrettable actions taken by the Cheney administration that books could be
written about them, and have been. My purpose here is to recount some of
the ones that illustrate Cheney’s attempt to grab power from Congress, ignore
the Constitution, favor corporations, retrench on New Deal advances for
the middle class and the poor, and bring America closer to a “proto-fascist
state” headed by the president as a dictator/king.
These are strong
accusations. They deserve a review to show how my chapter’s theme is supported
by the Cheney administration. But before doing that, let’s ask how America fared during the
Cheney Era.
Cheney “Results”
During the Cheney
administration the national debt almost doubled, from $6 trillion to $9.5
trillion in early 2008 (about $100,000 per family). This will take decades
to pay off, assuming the American economy does not slip into another recession.
This burden will fall on future generations, maybe some not yet born.
A report in 2005
stated that Americans spent $42 billion more than they earned, and the most
recent report from the Commerce Department found savings rates at a negative
one percent, the lowest since the Great Depression, and down from 11 percent
after WWII. An update for 2008 would show a far worse savings rate. Only
four times have savings rates fallen so low: The other two were during the
Great Depression when a quarter of the workforce was unemployed and Americans
spent their savings for essentials such as food and rent.
E. Manning writes
the following (TNTalk.). “Many millions of jobs … have been moved to distant
locations outside the U.S. where labor costs
are significantly reduced. …Multiple times, Congressional measures have been
considered to restrain the federal government from granting contracts to
companies that send work overseas. The Bush Administration has consistently
opposed the legislation. The administration has continually supported tax
incentives to outsource jobs overseas. … The government has worked as a
partner to stem public scrutiny of job outsourcing with no federal reporting
requirements for outsourced jobs. Multinational corporate America is fearful over
a public backlash of opinion that would prevent them from exercising complete
autonomy over their corporate work forces. … The Brookings Institute [estimates]
that outsourced re-employed workers recover 47 cents of every dollar that
they used to earn.”
Ohio has seen a loss
of 209,000 jobs, or 3.7%, during the first 7 years of the Cheney administration.
The US job loss during
this period was 19.5%. 61% of Americans fear job loss due to outsourcing.
In 2004 the head of the president’s Council of Economic Advisors, Gregory
Mankiw, described the advantages to US companies for outsourcing jobs overseas.
Mankiw said “When a good or service is produced more cheaply abroad, it makes
more sense to import it than make or provide it domestically.” “…outsourcing
is just a new way of doing international trade. …that’s a good thing.” Federal
Reserve Chairman Alan Greenspan defended Mankiw.
A typical worker
in a Chinese factory earns ~50 cents per hour, without the protection of
workplace safety regulations or limits on overtime. Chinese factories also
have fewer environmental laws to deal with. How can American workers compete
with workers in Communist China? From the standpoint of a manufacturing company
it would be a good thing to use slave labor, or maybe invent a machine that
could reduce their labor costs to zero by dispensing entirely with workers.
It would make sense for a company to adopt such a cost-saving change, but
only if very few other companies did the same. After all, if there were no
jobs for workers anywhere, who would buy the products? The “free market” left
to itself cannot handle this situation; this is a role for governments, which,
after all, are supposed to represent the interests of all the people.
The trade deficit
in early 2006 was $817 billion, and rising each year. About a third of this
imbalance was due to trade with China. Foreign interests
hold about $2 trillion. More than half of US Treasury bonds are bought by
foreigners. US companies and infrastructure
are being purchased by foreigners with the American dollars that just keep
accumulating in their bank accounts.
The unemployment
rate is rising as I write. It now stands at 6.1% (mid-2008). When discouraged
former workers are included the rate is ~ 10.3%. For reference, during the
Great Depression it reached 25%. Because of American worker wage losses the
credit industry generates over $30 billion in annual profits.
David Walker, the
US Comptroller General
(who audits the federal budget books)
has been traveling throughout the country warning Americans
about the dire economic situation. Here are some comments that I found on
the internet (apologies for not keeping track of the sources). “Foreign governments
and investors now hold fully half of the United States' total outstanding
debt, making Washington susceptible to a new form of geopolitical conflict”
“Japan and China are America's two biggest lenders. Great Britain is third, followed
by a bloc of oil-producing states including Iran, Kuwait, Saudi Arabia and Libya.” “So in trade
and military disputes, China, as America's No. 2 lender,
holds considerable influence.” “Foreign lenders, Bixby notes, can demand
conditions - or threaten to stop buying U.S. Treasury securities, or even
dump their existing holdings outright. To lure other buyers of Washington's debt, U.S. interest rates
would then have to rise sharply, throttling the nation's economy.” "It means
foreigners have more leverage on us and we have less leverage on them,"
Walker said. "You have
to pay attention to your bankers." “If the US could no longer
borrow as it has been doing, interests rates would
skyrocket, home values would plummet, people would lose their jobs, and
government services would have to be cut drastically. Shantytowns could spring
up, making America look more like
Haiti and South America.” "I'm sure that
people during the Roman Empire never thought that
Rome would fall," Walker said in an interview
last week. "I don't think we should assume that we are too big to fail." “All…agree it's a fantasy to argue that the U.S. can grow its way
out of its debt,” Bixby said. "The economy would have to grow at an implausible
rate forever."
Critical Review
of the Cheney Era
During the Cheney
Era government power flowed to the presidency at the expense of other parts
of the Executive branch and also at the expense of a “rubber stamp” Congress.
Many laws were passed that appear to have been drafted by corporate lobbyists,
and may have been coordinated by Cheney’s office. The effect of the laws
has been to loosen regulations affecting corporations, lower corporation taxes
and allow them to exploit cheap labor in foreign countries. The Cheney administration’s
neglect of enforcing employer sanctions for hiring illegal aliens allows
corporations to exploit workers by paying lower wages and neglecting safety
regulations. Laws that encourage globalization led to job outsourcing to
countries with cheaper labor (which began with Clinton’s NAFTA, to be
fair to Cheney). Tax law changes favored wealthy individuals and punished
poor and middle class wage earners. Credit card purchasing allowed strapped
wage earners to stay afloat, but because the credit industry is allowed to
charge usurious interest rates many people are driven to bankruptcy. Health
insurance continued to rise much faster than other family budget items,
and this may be due to an unwillingness to resist laws drafted by pharmaceutical
industry lobbyists that reduce competition. America’s energy policy
continues to be based on oil, with minimal funding of alternative and renewable
energy technologies. The median family income (with an ever-growing number
of two wage earners) has decreased slightly during the Cheney Era, while
the top 1% have enjoyed unprecedented income increases.
As an aside, Jamie
Johnson produced a movie “The One Percent” that documents attitudes of the
wealthy. He is descended from the founder of Johnson & Johnson, and belongs
to the wealthy class himself, so he had access to America’s wealthiest people
for the movie. In a recent web post (http://www.huffingtonpost.com/jamie-joh)
he remarks that some of the wealthiest are looking forward to a recession
for selfish reasons! It will result in a “thinning of the aristocratic ranks”
and lower prices for luxury goods that they have had to pay more for in recent
years. In other words, the ultra-rich resent the very-rich, and the poor
others can just go fly a kite! Johnson writes “…under the threat of hard
times the mega-wealthy aren't feeling a greater responsibility to reflect
upon the problems surrounding the growing wealth gap; they are, in fact, trying
to fatten their wallets and further insulate their lifestyles. I had hoped
that foreboding economic circumstances would have caused the ultra-rich to
think not just of themselves and increasing their own personal affluence.
Unfortunately, however, too many of them lack concern and without this concern,
the divisive imbalance will only worsen with recession.” I highly recommend
viewing the above URL for it contains more damning evidence than I could
include here. This rare insight into how the very wealthy think, and how little
regard some of them have for ordinary people, helps explain why corporate
executives can do some of the shameful things they do. It may also account
for the radical core beliefs of the hard-core Republican conservatives, like
Cheney.
Over and over, during
the Cheney Era, the wealthy have prospered while the poor and middle class
have suffered. This cannot be an accident; it has to be the result of policy
changes that are part of a long-term project to return America to a less democratic,
more authoritarian form of governance by the wealthy elite.
When America declared independence
from England 232 years ago there
were many who preferred to be ruled by King George III. These “royalists”
had a resurgence during the Gilded Age of the
19th Century. And another resurgence during
the Roaring Twenties. Their most recent resurgence occurred during
the Reagan to Cheney era.
Government has become
a better partner with business than at any time since 1929. The 9 years that
led to the Great Depression resemble the 25 years since Reagan became president.
The slowness of the present economic decline is due to the ability of the
rest of the world to loan money to America. But America is now so deep
in debt that the outcomes for these two periods may eventually be the same.
The American government
is no longer “Of the People, By the People and For the People.” Instead,
the government is “Of Corporations, By Corporations and For Corporations.”
America has quietly created
its own version of a Proto-Fascist State!
Let’s review Mussolini’s
definition of Fascism to see how accurate my indictment is: “…a
system of government that exercises a dictatorship of the extreme right,
typically through the merging of state and business leadership, together with
belligerent nationalism.” The word “dictatorship”
is partially fulfilled because Cheney uses his figurehead president to
issue signing statements that overrule Congress. The “merging of state
and business leadership” is almost completely fulfilled, thanks to the
common practice of lobbyists drafting laws that elected congressmen are
expected to pass (in exchange for campaign contributions). And “belligerent
nationalism” was demonstrated by the way America installed a new regime
in Iran in the 1970’s; by the American CIA assisted assassination of Chile’s
socialist president Allende in order to pave the way for murderer Pinochet
to take power; and by America’s unwarranted invasion of Iraq with dubious
provocation. I think this demonstrates “belligerent nationalism.”
America’s evolution to
a proto-fascist state may have gone too far to be reversed. Once again in
the history of humans, power wins! The strong have figured out a way to steal
from the poor, and they’ve done it so quietly that the poor don’t even realize
that they’ve been victimized. So naturally the poor are unaware of who’s
been doing the stealing. The victim’s pockets were picked so expertly that
with their clueless vote the pick-pocketers continue to enjoy everlasting
opportunities for continuing their scam.
Will America
Avoid a Great Depression?
The question to
ask now is “Will America descend into another
Great Depression?” Let’s review the similarities of the 1920’s and the 25-year
Reagan/Cheney era:
● During the 1920’s, businesses achieved 32% productivity
gains but wages increased only 8%. During the Reagan/Cheney era business
productivity rose 40% but for 72% of workers wages declined. A new force
came into play during the Reagan/Cheney era: corporations fired American
workers and hired cheap foreign ones.
● During the 1920’s, median savings dwindled to negative
values just before the Crash. During the Reagan/Cheney era median savings
dwindled from positive values to
-1%. In 2005 the poorest 10% of families had a negative net worth.
● During the 1920’s, installment credit purchasing increased
as a solution to falling purchases and filling warehouses. During the Reagan/Cheney
era the credit industry boomed for the same reasons, reaching $30 billion
per year.
● During the 1920’s, Republicans controlled government
and they empowered their “rule” through corrupt and immoral stratagems. During
the Reagan/Cheney era the same occurred.
● During the 1920’s, the income gap between rich and poor
grew to the staggering state in which 0.1% of people controlled the same
wealth as the lowest 42%. I don’t have the 2008 statistic for the top 0.1%,
but in 2008 the top 1% controlled about half of America’s wealth (Jamie
Johnson, 2008). During the 21 years from 1984 to 2005 the top 2% of American
families doubled their wealth while the bottom 25% lost wealth (corrected
for inflation). During the Reagan/Cheney era, the ratio of a Fortune 500 CEO earnings to the average worker’s
earnings changed from 40:1 to 424:1 (Allan Ornstein, 2008). In 2006 the
top 1% earned 20.3% of all income.
● During the 1920’s, “…the Revenue Act of 1926 … reduced
federal income and inheritance taxes dramatically.” In just 5 years
Reagan lowered taxes for the wealthy from 70% to 28%, while raising rates
at the low end of the income scale.
Had enough? If you
want to be scared some more, go back and re-read the section on the Roaring
Twenties, and note the many similarities to the Reagan/Cheney era.
Theoretical
Extrapolation of Current Trends
This penultimate
section of the chapter is a valiant if not foolhardy attempt to predict the
future of America
using the two theories that underpin the chapter.
Theory #1 states
that during times of war, or times of extreme hardship that resemble war,
people are likely to sacrifice themselves on behalf of the over-riding goal
of tribal survival. For maybe 50,000 years this meant that at the onset of
inter-tribal conflict ordinary men would become patriotic warriors and everyone
else would support them with whatever patriotic acts strengthened the tribe.
The theory also states that in times of peace tribal survival is not threatened,
which releases people from patriotic duty. With the breakout of peace people
can be expected to act in ways that I have described as “nasty and selfish.”
This includes a form of greed accompanied by an unwillingness to share one’s
good fortune with those who made it possible, as well as a mocking indifference
to the poverty and suffering of others. Everyone who can will aspire to
wealth, and those who succeed will continue to
exploit those who have not succeeded.
Theory #2 states
that the last 12,000 years of conflict between super-tribes, that can decimate
the losing tribe, evolution has favored people who are programmed to prefer
a form of governance in which decisions are made from the top by a strong
leader. Nevertheless, there is also a minority (of pre-Holocene “throwbacks”)
who are inclined to prefer a more democratic form of governance in which
the people make decisions that regulate how everyone is to behave, regardless
of their strength.
Maybe these two
theories can be combined into the following prediction: During war (or times
of extreme hardship) people demand strong leadership for coordinating the
efforts of everyone else, who in turn are all deemed essential for the group
effort to survive. During times of peace people want to be left alone to
pursue their own aspirations to prosper. But those who prosper the most (achieve
wealth) take advantage of their power to exploit and subdue the others. Since
most people are susceptible to being followers (because following during
war is essential) they may follow when it is not in their best interest to
follow. The danger of this is especially great when a powerful leader is
very clever at redefining which social condition is operative. This can lead
to an unwanted leadership that neglects the aspirations of everyone who lacks
power.
The illegitimate
leader situation is self-limiting, because a society requires the productive
labors of that “man with the hoe” and if his hoe is taken from him he cannot
produce. Every empire is destined to collapse following corrupt leadership.
After the collapse the productive peasant reclaims control of his life and
it is his responsibility to figure out how to prosper, and in the process
maybe create another empire. This implies that history can be viewed as a
pendulum that swings back and forth between the two states. The pace of the
pendulum’s swing may vary greatly.
Let’s review how
the pendulum has swung during America’s
232 years as a country. It will be convenient to invent terms for referring
to the two extreme states of the pendulum. How about
“producer” and “parasite.”
Starting with the
War of Independence, which is “producer,” we see a slow swing to the “parasite”
state called the “Gilded Age” (1877-1893). During the “Progressive Era” (1896-1920)
the pendulum swung back to “producer.” The 1920’s saw a fast swing to “parasite,”
culminating in 1929. The New Deal era of FDR saw a swing back to “producer”
– which endured until Nixon’s election (1968). Slowly the pendulum swung
in the “parasite” direction until it got a big boost from Reagan, which caused
the parasitic movement to accelerate. With Cheney the pendulum has swung
to a “parasite” state that is about as far as it can go. What, we now ask,
what will the pendulum do next?
At the risk of overusing
a metaphor, I allege that there’s no guarantee that the pendulum will keep
swinging. When it goes too far in the “parasite” direction it may be “caught”
and held there.
You may have thought
that I was going to predict that the pendulum would soon swing in the “producer”
direction. This would be good, but for this to happen the corrupt leadership
would have to “return the hoe to that man who wants his hoe.” If the wealthy
Americans refuse to give that man his hoe, America
could become a fascist state!
This happened in
Italy
and Germany
75 years ago, and it happened in Japan
somewhat earlier.
During the Reagan/Cheney
interval America’s
wealthiest people have prospered from productive labors that have shifted
ever-so-slowly, but irreversibly, to workers in third world countries.
The American worker has had his hoe taken from him by the parasites, and
those parasites have given the hoe to workers overseas.
But the parasites
“made a deal with the devil” as they did this. The producers overseas have
started to own America.
It’s more accurate to say that the overseas governments have begun to own
America.
Those overseas governments are also prone to exploit their workers, and
when American corporations wanted the labors of workers in those overseas
countries the governments saw an opportunity for improving their wealth,
so they allowed the corporations to locate their factories there. America
now has “its hands tied.” It cannot object to anything our creditors want.
If China
wants to take over Taiwan,
they can do it with impunity. If Saudi
Arabia wants to buy controlling
interest in profitable American companies, the laws will be changed to allow
that rather than risk financial ruin.
Little by little
America
will become fractured into pieces that resemble colonies belonging to many
other countries. China
will own mines that produce the minerals they need, they will buy farms
that grow the wheat, and the cattle ranches that used to provide meat to
Americans will ship the meat to China.
America
will become a source of resources that will flow to our creditor nations,
and Americans will be working for foreign employers. It will be a reverse
of the present situation where Chinese workers make Nike sneakers for export
to America.
Order will have
to be maintained in America
for our colony status to remain valuable to our creditor nations, and this
may be done by the American parasites using a fascist fist. Cheney is suited
for this job, but as he retires there will be other Cheneys eager to exercise
fascist discipline.
The grim portrait
of America’s
future may not happen. I hope it doesn’t happen. But it may happen!
Prospectus
for America
I mentioned earlier
that I was not going to vote in the 2008 elections. This is because I believe
America’s
fate is sealed, and nothing can turn around the inevitable slide to fascism.
McCain/Palin might
actually be good for Planet Earth! The sooner America
slips into depression, the sooner our Earth will be relieved of a profligate
polluter and destroyer of nature.
It’s frustrating
to see things about America
the way I do. Sometimes, when I feel a pang of optimism, I hope that I’m
wrong. But I always come back to this unhappy picture. Imagine standing at
the bow of the Titanic, seeing an iceberg, shouting warnings to the others
on the ship, but no one listens! That’s how I feel.
American voters
have lost the right for a winning place in the world. When voters get what
they want, they deserve what they get – “good and hard” as Mencken would
say. Long-term prosperity has to be earned, and the pursuit of short-term
prosperity by taking short-cuts has a deservedly uncomfortable end point.
If I were younger, I would try to relocate to New
Zealand. But as a retiree,
nestled on a small acreage in the Arizona
countryside, I shall sit back to watch the unraveling of a country that
I once loved.
─────────────────────────────────
CHAPTER 20
─────────────────────────────────
THE FALL OF CIVILIZATIONS
‑ PART II
DATING THE DEMISE
OF HUMANITY
Introduction
This chapter relates
world population data with science and technology innovations and arrives
at a "per capita rate of innovation" graph. The "per capita rate of innovation"
shows two peaks, one starting during the Golden era of Greece and the other
starting during the Renaissance and peaking at the end of the 19th Century.
A range of dates for the demise of humanity is calculated on the very speculative
principle that there's a 50% chance that we now find ourselves between
the 25th and 75th percentiles of the sequence of the birth dates of all
humans who shall ever be born. In 1990 I wrote a brief version of this essay,
dealing specifically with the statistical argument for inferring that the
demise of humanity was imminent; it appeared in an unpublished book, Essays
From Another Paradigm. The present chapter
is adapted from a 1993 expanded essay on the same subject.
Population Versus Time
Table 1 is a compilation
from many sources of the world's population for 26 epochs. The original literature almost never provides uncertainties,
but if scatter is any guide the uncertainties range from 3% during this century,
to ±3 dB (+100/‑50%) at 8000 BC, and ±5 dB at 100,000 BC.
Table 1
Year Pop’n Year Pop’n Year Pop’n
[AD] [millions]
[AD] [millions] [AD] [millions]
‑100,000 0.5
1500 440 1950 2530
‑50,000 1 1600 470 1960 3000
‑18,000 3 1650 545 1970 3600
‑8,000 9 1700 600 1980 4400
‑7500 10 1750 725 1990 5300
‑3000 30 1800 907 2000 (6380)
‑1000 110 1830
1000 2010 (7300)
0 200 1900 1600 2025 (8500)
1000 340 1930 2000 2038 (8500)
A 10th order polynomial fit
to the relationship of "log of population" versus "log of time" is given
in Appendix D. It has been used to perform integrations from the distant past
to dates of interest. The following figure plots the tabulated data (symbols)
and the 10th‑order fit (trace).
Figure 20.01. World
population versus time, using a special Log scale for time. The trace is a 10th order polynomial fit, used to assist
in later calculations.
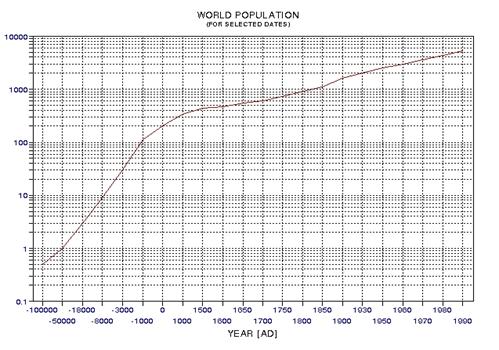
Figure 20.02. Adopted
world population, with arbitrary choice of year for x‑axis representation.
Birth and Survival Rates
Before proceeding to the calculation
of the integrated number of human live births and adults, it is necessary
to address the issue of birth and survival rates. The simplest method for
calculating the integral of population from some arbitrary start time to
x‑axis time is to multiply "crude birth rate" times "population" times "time
interval." I've adopted a crude birth rate table that starts at 45 births
per thousand at 100,000 BC, and decreases monotonically to 26 births per thousand
in 1993. It has been established that the main decrease started at approximately
the time of World War II, when it had a value of 38 births per thousand.
Not all babies live to adulthood. Throughout the world prior to the 18th
Century approximately 25% of babies survived to adulthood (taken to be the
age when reproduction begins, about age 18 in primitive societies, and age
13 in developed world societies). In other words, in the natural order of
things approximately 3/4 of all newborns are destined to die before adulthood!
Since the 18th Century the developed world has achieved a much better survival
rate, approximately 95% (versus 25%). But still, the undeveloped world (about
71% of the world's population) has survival rates of approximately 30 to
35%. The adopted world average survival rate conforms to estimates of the
fraction of the world's population that is "undeveloped" versus "developed."
The adopted birth and survival rates are shown in the following figure.
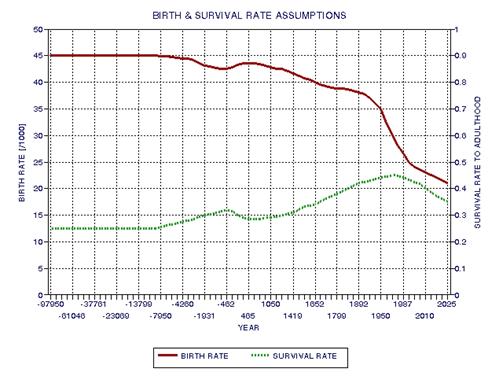
Figure 20.03. Adopted crude birth rate
(solid) and survival rate (dashed).
Integrated Population Versus Time
The previous graphs illustrate
time interval averages for population, birth rate and survival rate. These
are combined to calculate the integrated number of births from 100,000 BC
to x‑axis time. In the following figure the upper trace is labeled "live
births." Thus, this trace is the total number of live births from 100,000
BC to x‑axis time. Note that the x‑axis is neither
linear nor logarithmic, but corresponds to dates in the original population
data, above.
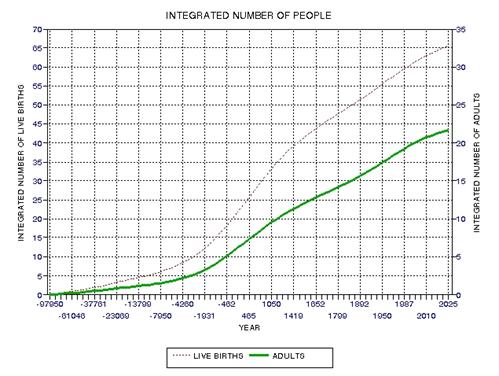
Figure 20.04. Integrated number of live
births (dotted) and integrated number of humans reaching adulthood (solid).
Note the solid trace, the
integral of adults who have inhabited the earth from 100,000 BC to x‑axis
time. To calculate this it was necessary to use the estimated survival rate
versus time (the lower trace in Fig. 20.03). The number of "adults" that
have inhabited the world is about 33% of the number of all humans born. For
the epoch of these calculations, 1993, the total number of "live births"
was 60.3 billion, and the total number of adults who have ever lived (to
1993) is 19.6 billion.
As an aside, Fig. 20.05 is
a plot of the D/L Ratio, defined as the ratio of dead to living. This parameter
was apparently treated by Asimov (reference unavailable).
At the time of this writing
(2001) the D/L Ratio is about 8.6. Figure 20.06 shows how the ratio "people
alive at date" to "total births to date" has varied over time.
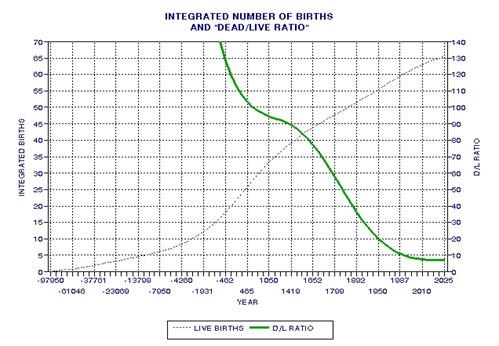
Figure 20.05. Ratio
of dead to living (solid trace) versus time.
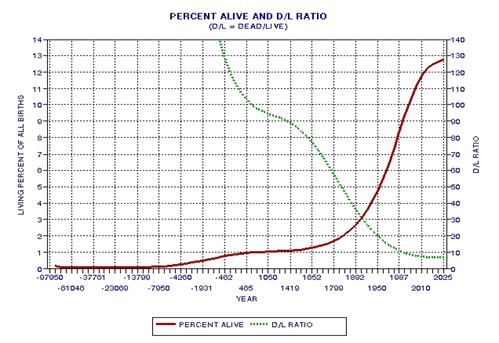
Figure 20.06. Percentage
of "all births" represented by "people alive."
In the year 2006, when 6.8
billion people are supposedly alive; they constitute 11% of all people who have ever
been born.
The following figure is an
alternate presentation of the data in Fig. 20.04, with a rescaling of the
y‑axis so that in 1993 the integrated number of people is 100%.
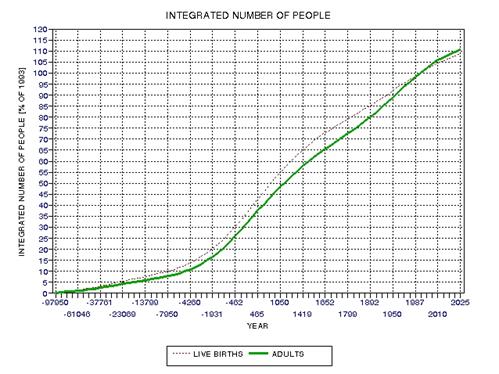
Figure 20.07. Integrated number of human
live births and humans reaching adulthood, normalized so that 1993 has 100%
of the integrated numbers.
The above figure plots the
"integrated number of people" as a percentage of the 1993 numbers. The
"live births" and "adults" traces cross at 1993, by definition. These traces
can be used to define what I shall call the "Humanity Time Scale." Which
of these traces should be used? The "live births" trace has fewer assumptions;
just the population versus time and the birth rate versus time, both of which
are well established. The "adults" trace may be more appropriate for what
we are going to do with the Humanity Time Scale as it reflects the number
of humans who have lived long enough to think about the world, and contribute
to it's irreversible legacy of innovations. The
weak part of the argument for adopting the "adults" trace is that it depends
on survival rate, which is an assumed parameter. It is less well‑established
than the other two properties. The halfway points (the 50% level) for the
two traces are at 834 AD and 1118 AD, for "live births" and "adults."
The following figure is a plot
of "% of adults before date" versus year for a set of arbitrarily chosen integer
dates.
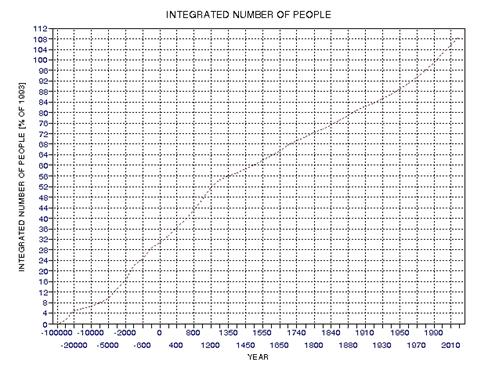
Figure 20.08. Integrated number of human
adults born before (arbitrarily selected) x‑axis years.
It is slightly easier to use
this graph to determine dates before which specified percentages of all
human adults were born. For example, 80% of adults lived prior to the year
1891 AD, and 82% of adults lived before 1908 AD. Thus, 1891 to 1908 AD is
a "2% of adults" interval (corresponding to 80 to 82% of adults). There are
50 such 2%‑intervals prior to 1993, and each has corresponding beginning
and ending dates.
Innovation Data
“Asimov's Chronology
of Science and Discovery" (198?, 1994) has
been analyzed to determine how many innovations belong to each of the 2%‑intervals.
Asimov's list has 1478 entries, from 4 million BC to 1991. For the time span
100,000 BC to the present, there are 1474 items. A histogram was created
showing the number of items for each 2% date interval. For example, for the
2% date interval 1891 to 1908 AD, there were 120 citations in Asimov's list.
As there are 2% of 19.6 billion adults during each 2% interval, or 392 million
adults, the number of innovations per billion people can be calculated by
dividing the number of citations by 0.392. The results of this conversion
are presented in the following figure.
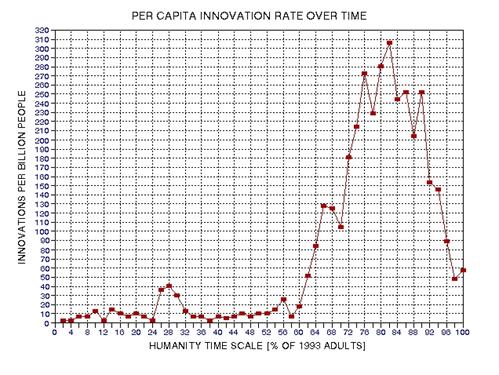
Figure 20.09. Number
of innovations per billion adults for each 2% interval of the Humanity Timescale.
The first peak, at 28%, the
2% interval of 26% to 28%, corresponds to 500 BC to 290 BC. The minimum at
38% corresponds to the dates 390 AD to 500 AD. The abrupt rise after 60%
corresponds to the mid‑15th Century, which is when the Renaissance began
(1453 AD). The peak at 82% (corresponding to the 80 to 82% time interval
cited above) is for the period 1891 to 1908 AD. The steady decline since
1908 has progressed to a level corresponding to that of the 16th Century.
Weighted Average Innovation
Rate
About 96% of Asimov's science
and discovery citations belong to a category that requires formal education,
by my cursory review. It is thus natural to ask how many "literate" people
there have been over time, and how does the innovation rate look when it
is normalized to the relative numbers of literate people? Better, how does
the innovation rate look when it is normalized using a 96% weight for the
literate population and a 4% weight for the illiterate population?
To normalize the innovation
rate traces to the population of literate adults it is necessary to adopt
literacy rates over time. I have chosen to do this on a region‑by‑region
basis, since literacy commences at different times in different world regions.
It is also necessary to estimate regional population traces. I have chosen
9 world regions for this task. Figure 20.10 shows the population of 5 regions
(the most populace), and Fig. 20.11 shows the population of the remaining
4 regions.
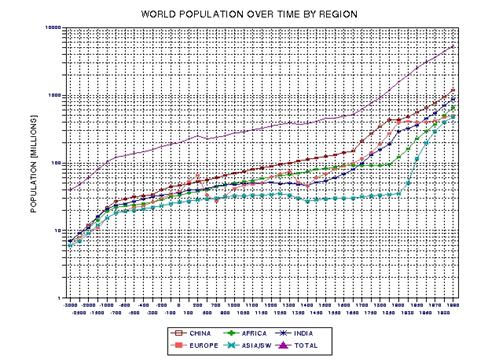
Figure 20.10. Population
breakdown for 5 regions and their total.
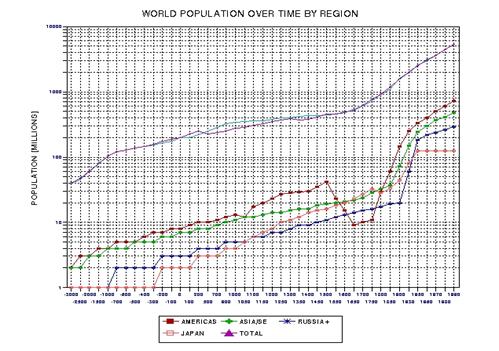
Figure 20.11. Population
breakdown for another 5 world regions, and their total.
Notice that in Fig. 20.10
Europe experienced two population peaks before the Renaissance: in 200 AD
and 1300 AD. There are population collapses after each peak. The first collapse
must have something to do with the inability of urban centers to support
large populations (the population of Rome fell dramatically, for instance),
while the second collapse was produced by the scourges of the Black Death.
In Fig. 20.11 there is one (documented) population collapse, starting in
1500 AD, caused by diseases brought to the New World by European explorers and
settlers. The population rise starting in 1750 is due to massive migrations
of Europeans.
It was not possible to find
literacy rates for all these regions for the times of interest. After the
suggestion of Dr. Kevin Pang, I adopted the procedure of estimating literacy
rate by assuming that most urban populations are mostly literate while
most of the rural populations are illiterate, at least until recent times.
Urban and rural statistics are easier to estimate, so this procedure can
be used for more regions and can be extended back in time to the adoption
of writing in each region. In constructing these tables it was assumed that
approximately 50% of the pre‑15th Century urban population was literate,
and approximately 1% of the rural population was literate. After 1500 AD
a gradual increase in the two literacy rates are adopted, ending with a
present day 90% and 40% (weighted average of all regions).
Other minor adjustments were
made as an attempt to represent "realism." For example,
for the Americas the literacy rate was allowed
to climb from zero during the first Century AD, when the Mayan “civilization”
is thought to have adopted writing. The Americas literacy rate remained at
low levels during the pre‑Columbian era, and rose
rapidly during the European immigration. Similar "origins" of literacy are
attributed to China in the 17th Century BC, and "Europe" (actually Mesopotamia") during the 4th Millennium BC. Regional literacy rates were
combined with regional populations to produce a global literacy rate and
total number of literate adults, which is shown in Fig. 20.12.
Figure 20.13 is innovation
rate per literate adult. It is a renormalization of Fig. 20.09, using the
global literacy rate as a normalizing factor; so it thereby retains the property
of showing how many innovations were produced per million literate adults
who lived during the “equal increment of adults” intervals.
It is remarkable that after
the classical Greek period the rate of innovations is level at about 50 per
million literate adults until well into the 19th Century. This could be
the source of interesting speculation, but for now I will defer. The pre‑Greek
times produced innovation rates comparable to those of the Greek era, but
this feature is less robust for several reasons: 1) there are fewer innovations
in the numerator, and 2) there is great uncertainty in estimating (or even
defining) literacy during this time.
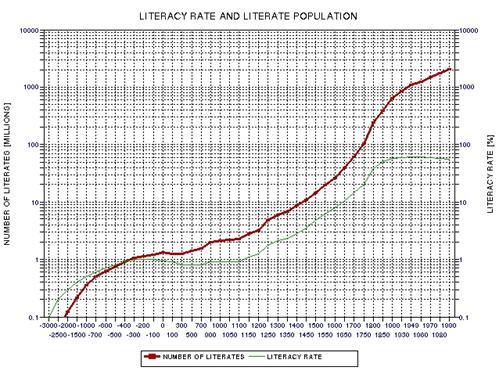
Figure 20.12. Estimated
global literacy rate and total number of literate adults versus time.
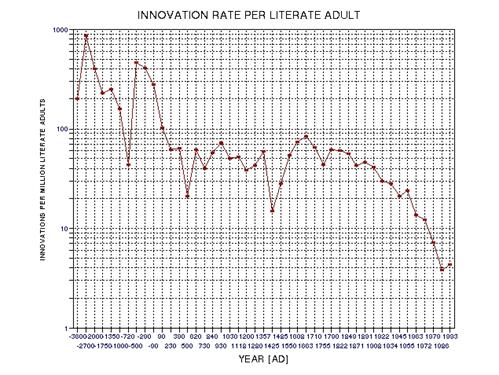
Figure 20.13. Innovation
rate per literate adult.
The drop in innovation rate
since 1800 is attributable to two equally important factors: 1) a population
that rose by a factor of 5.5, and 2) literacy rate grew by a factor of
3.8. Since both factors move the innovation rate trace in the same direction,
a factor of 21 decrease is predicted due to these
two considerations alone (while a drop of 15 to one is observed).
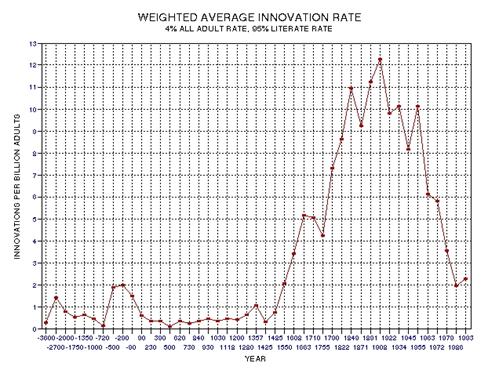
Figure 20.14. Innovation
rate per billion population, weighted average of rates for literate and all
adults.
This figure is a plot of the
innovation rate using the weighted average of 4% for illiterates and 96%
for literates. This trace is based on the concept that the literate person
is 24 times as likely (96/4 = 24) to produce an innovation (that Asimov would
include in his list) compared to the illiterate person. This presentation
is the "fairest" way that I can think of for representing innovation rate
using Asimov's compilation as the measure for significant innovations.
The Two Major Peaks in Innovation Rate
There are still two peaks in
Fig. 20.14, as there were in Fig. 20.09. The classical Greek peak in relation
to the 19th Century peak is 13% in Fig. 20.09, and 17% in Fig. 20.14. Normalizing
by a weighted average of literate people and illiterate people's overall
productivity did not significantly change the relative appearance of the two
versions. The Greek peak endures for about 4 centuries, from 500 BC to 90
BC. The 19th Century peak occurs between 1550 AD and 1993 AD, approximately,
which is about 4.5 centuries long. Thus, the durations are approximately
the same in terms of normal, calendar time, being 4 or 5 centuries. I will
refer to this most recent peak as the Renaissance/Enlightenment innovation
peak.
There is another similarity
between the Greek and Renaissance/Enlightenment peaks. They are both accompanied
by an increasing population, and the Greek population rise reaches a maximum
some centuries later. The Greek infusion of new ideas was exploited by the
Romans, who made it possible for populations to increase until a collapse
after 200 AD. The population maximum occurred 5 centuries after the innovation
peak. Figure 20.15 illustrates this.
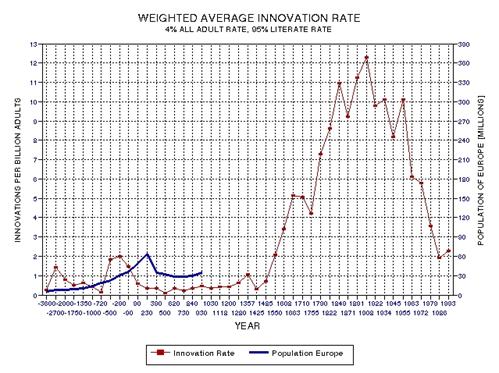
Figure 20.15. European population in
relation to global weighted‑average innovation rate, showing that the "Greek"
innovation peak is followed 5 centuries later by a "Roman" population peak.
Figure 20.16 shows a 1400‑year
expanded portion of the previous figure, centered on the Greek innovation
peak. The Roman population peak follows the Greek innovation peak by 4 to
6 centuries.
Figure 20.17 shows another
1400‑year period, centered on the Renaissance/ Enlightenment innovation peak.
Clearly, this dynamic cycle is still unfolding, and we alive today are naturally
interested in its outcome.
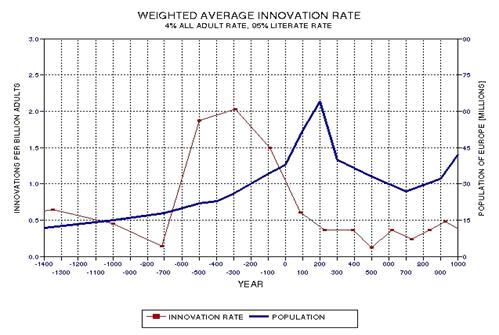
Figure 20.16. A 1400‑year expanded portion
of the previous figure, centered on the Greek innovation peak.
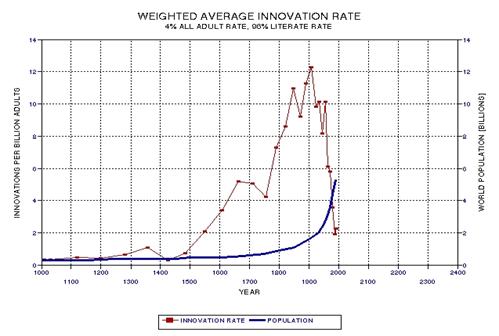
Figure 20.17. Another
1400‑year period, but this time centered on the Renaissance innovation peak.
It is inevitable that the
still-unfolding Renaissance/Enlightenment innovation peak will be followed
by a population peak, and I conjecture that its timing will be similar to
the timing of the Greek innovation and Roman population peaks. We do not
know the future, but some population projections resemble the plot in the
next figure, with a population peak in ~2200 AD, and a collapse afterwards.
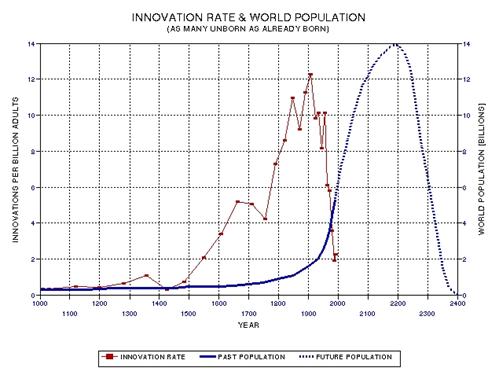
Figure 20.18. The
same Renaissance 1400‑year peak period, but with a future population trace,
showing a population peak aafter the innovation peak.
Actually, this particular
future population curve is a special one, for which I shall present an argument
in the next section. Note, for now, that the population peak occurs only
3 centuries after the innovation peak, whereas the Roman population peak
followed the Greek innovation peak about 5 centuries. By analogy, the currently
unfolding population explosion in the undeveloped world owes its existence
to the Renaissance/Enlightenment innovation peak at the end of the 19th Century.
It is also interesting that
for both pairs of innovation/population peaks, the innovations and population
growth occurred in different parts of the world. The spread of technology
from the site of its origin allows other populations to grow almost as surely
as it allows the innovating population to grow. This is reminiscent of the
old saying: "When the table is set, uninvited guests
appear."
Random Location Principle and
Forecasting the Future Population Crash Date
It is perhaps important to
put the upcoming population crash scenario to the test of what I shall refer
to as the Random Location Principle. After I performed the analysis presented
here I learned that the subject had been discussed in a late 1980’s publication
and was referred to as the “Anthropic Principle” (erroneously, I think).
The Random Location Principle states that "things chosen at random are located
at random locations." This innocent sounding statement is not trivial. It
can have the most unexpected and profound conclusions, as I will endeavor
to illustrate.
Before applying the Random
Location Principle (RLP) to the population crash question, let us consider
a simpler example that illustrates the RLP concept. Consider the entire sequence
of Edsel cars built. Each car has an identification number, thus allowing
for the placement of each Edsel in a sequence of all Edsel cars. Assume
for the moment that we don't know how many Edsels were manufactured, and
let's try to think of a way to estimate how many were manufactured by some
simple observational means. Suppose we went to the junk yard and asked to
see an Edsel. Assuming we found one, we could read the identification number
and (somehow) deduce that it was Edsel #4000 (the 4000th Edsel manufactured).
Would this information tell us anything about the total number manufactured?
Yes, sampling theory says that if we have one sample from the entire sequence, and if it is chosen at random, then if we
double the number in the sequence we'll arrive at an estimate of the total
number in the sequence. In other words, doubling 4000 gives 8000, which is a crude estimate of the length of the entire sequence.
Sampling theory goes further,
and states that we can estimate the accuracy of our estimate. Namely, we
can assume that a sample chosen at random has a 50% probability of being
within the 25th and 75th percentile of the entire sequence. If 4000 were near
the 25th percentile, then the sequence length would be 4 times 4000, or 12,000.
If 4000 were near the 75th percentile, the sequence length would be 4000
* 1.333, or 5300. So, with just one random sample, the number 4000 in the
sequence, we could infer that there's a 50% probability that the entire sequence
length is between 5300 and 12,000. Moreover, there's a 25% probability that
the entire sequence length is less than 5300, and another 25% probability
that it is greater than 12,000.
Now we’re ready to apply this
principle to the human sequence. Assume every human birth is assigned a
sequence number. Let's delete people who fail to reach adulthood, so our
new sequence is for all people born who eventually become adults. The next
step is going to be difficult for most readers, but I want to try it. Imagine
that the future exists in some sense. It's like watching a billiards game
and having someone exclaim that while the balls are moving the future motion
of the balls is determined. Thus, after the balls are set in motion the unfolding
of future movements and impacts is determined. For physicists it is somewhat
straightforward to conceive of the universe as a giant billiards game, set
in motion by the Big Bang 13.7 billion years ago. So imagine, if you can,
that there is a real sequence of unborn people who will be added to those
already born, and that this sequence is somehow inherent in the present
conditions. If it helps, think of time as a fourth dimension, and the entirety
of the future is just as real as the entirety
of the past, and the NOW of our experience is just a 3‑dimensional plane
moving smoothly through the time dimension. If you can accept this concept,
then the rest is easy.
Each person is just one in
a long sequence of people comprising the entirety of Humanity. Few people
can expect to find themselves at a privileged location in this sequence;
rather, a person is justified in assuming that they are located at a "typical"
location in the sequence. For example, there's a 50% chance that you and
I are located between the 25th and 75th percentile along this sequence of
all humans. If we are near the 25th percentile, and since 19.6 billion adults
were born before us, we could say that another 58.8 billion adults remain
to be born (i.e., 3 x 19.6 = 58.8). Or, if we happen to be near the 75th percentile,
we could say that another 6.5 billion people remain to be born (i.e., 19.6
/ 3 = 6.5). In other words, there's a 50% chance that the number of humans
remaining to be born is between 6.5 billion and 58.8 billion. To convert
this to calendar dates, we need to experiment with future population curves
to find those which end with the required hypothesized number of future adult
births.
Consider the future population
trace in Fig. 20.18 that goes to zero in 2400 AD. Integrating
it to 2400 AD yields 35 billion new adults. If this is humanity's
destiny, then those born in 1993 would be at the 56% location in the entire
Humanity sequence. Or, those who were born in 1939, as I am, would be located
at the 49% location of the entire Humanity Birth Sequence. These locations
are definitely compatible with the Random Location Principle, and the population
projection that goes to zero in 2400 AD is an optimal candidate to consider,
since it places today's adults near the mid‑point location of the Humanity
Birth Sequence.
However, we are searching for
a population curve that has an integral of 6.5 billion new adults, and also
a curve with an integral of 58.8 billion. Through trial and error I have
found two curves that meet these requirements, and they are presented as
Fig. 20.19.
The curve with a population
collapse to zero in 2140 corresponds to the hypothesis that we are currently
near the 75% location in the Humanity Birth Sequence. The population collapsing
to zero at 2400 AD is a most likely scenario, and corresponds to our being
near the 50% location. And the right‑most curve, with a population collapse
to zero at 2600 AD, corresponds to our current location being near the 25%
location. There is a 50% chance that the collapse will occur between the
two extremes. Thus, by appealing to the Random Location Principle, we have
deduced a range of dates for the end of humanity!
The future population shapes
can be rearranged, provided areas are kept equal. Thus, the real population
curve is likely to have a small "tail." I would argue that after such a colossal
collapse the people surviving and living in the tail would be genetically
and culturally distinct from today's human. Following the example of Olaf
Stapledon, in Last and First Men (1931), humanity after the collapse
will enter a transition from a First Men phase to a Second Men phase. New
paradigms will define the new man.
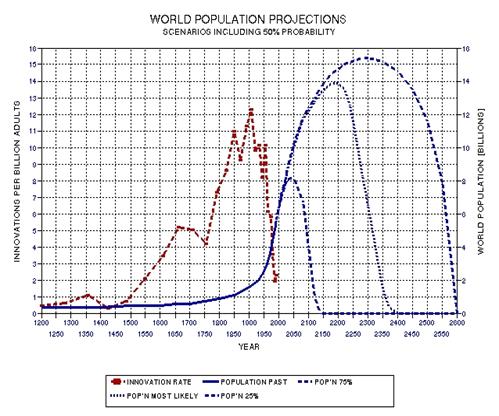
Figure 20.19. Three
future population scenarios, encompassing 50% of what is forecast by my usage
of the Random Location Principle. See text for disclaimers.
Final Humanity Time Scale
The following table lists
equivalences of "YearAD" and "Humanity Time Scale %." The table extends to
200%, corresponding to the "most likely" population crash date of 2400 AD.
The following figure is a
visual representation of the Humanity Time Scale described by the equations
(modified so that the year 2000 AD corresponds to the 100% point on the scale),
presented in Appendix D.
Caveat and Comment Concerning
Humanity's Collapse
The population collapse suggested
by the "Random Location Principle" is clearly speculative! Its claim for
consideration hinges on the applicability of the Random Location Principle
to the situation of a sentient being posing the question "where am I in the
immense stretch of humanity?" I suppose the conventional wisdom, if someone
representing it were pressed to respond to such a question, would say that
we are now close to the very beginning of this immense sequence, and that
humanity may exist forever. In addition, that person would say, when our sun
explodes in 5 or 6 billion years, humans will have migrated to other star
systems, and will have secured its rightful place as an immortal cosmic species.
HUMANITY TIMESCALE

Figure 20.20. Humanity
Time Scale. Left scale is for past, right is for past and future,
and assumes humanity (as we know it) ceases after 2400 AD. Equal intervals
along the vertical scale correspond to equal numbers of adults in the entire
sequence of births leading to adults
Well, that optimistic belief
requires a response to the following: “If humanity is going to endure for
another 6 billion years at something like its present population level and
lifespan, then isn’t it amazing that we are located at the 0.00006 % place
on the long sequence of human existences.” How likely is it that we are really
this close to the beginning of everything that will comprise the human story?
What a privileged position we would now have if this were true!
Additional Thoughts on the
Meaning of This Result
The Andromeda galaxy is moving
toward our Milky Way galaxy at 500,000 kilometer per hour, and the collision
date, assuming it's a direct hit, is approximately 3 billion years from
now (Science, January 7, 2000, p. 64). Speculation over consequences
has just begun, and initial thoughts are that a burst of new star formation
and supernova explosions might bathe the solar neighborhood with radiation,
photon and particle, that could pose a hazard to all Earthly life, or that
too many comets will be forced out of the Oort cloud and increase the rate
of climate disrupting impacts. I assert that Humanity may not survive the
present millennium, so "not to worry!" about things 3 billion years from
now!
If only such optimism as worrying
about hazards 5 or 6 billion years from now were warranted! Of course,
none of us know if this will be true. We must be content with speculation.
And mine is merely one, conceivable speculation.
It surprised me to discover
that for the past century the innovation rate has been decreasing. At first
I thought this must be due to an under‑representation of innovations from
the 20th Century. But the absolute number of innovations continues to increase
during the 20th Century. There's a simpler explanation. The innovations
are coming from slow‑growing populations of America, Europe, Australia, New Zealand and some Asian countries,
while the world's population can be attributed almost entirely to the undeveloped
countries. Thus, even though America and Europe, and parts of Asia, are producing an ever‑growing
number of innovations, and perhaps growing on a per capita basis, world
averages show an innovation rate decline.
The careful reader may have
wondered "The causes for a population rise following a spurt of innovation
are easy to imagine, but what could cause a decline? This subject is treated
in Chapters 11, 14 and 16.
It has just come to my attention
(March 16, 2000) that many people have independently stumbled upon the idea
for inferring the imminent demise of humanity, as we know it, using what
I referred to as the "Random Location Principle" ‑ but which apparently has
a generally accepted name, the "Doomsday Argument," and which is closely
associated with a related topic referred to as the "Anthropic Principle."
My original essay on this subject, “A New Estimate for the End of Humanity,”
appears in Chapter 7 of my 1990 book Essays From Another
Paradigm (self‑published, not for sale). This essay actually post‑dates
similar writings by others by a few years, but I wasn't aware of any of these
writings until about 1995. A good starting point for learning what others
have written about the subject can be found at: http://www.anthropic‑principle.com/profiles.html
One intriguing way to reconcile
the “Doomsday Argument” with a long human lifespan is to assert that a
"long individual human lifespan" is compatible with the Random Location
Principle and a very long human existence. If biotechnology affords some lucky
individuals the means for achieving immortality, they may come to dominate
world affairs and eventually extinguish the mortal sub‑species of humans.
Then, the number of humans ever born will have reached a final maximum number,
on the order of 2 or 3 times our present accumulation, and the Random Location
Principle viewpoint will remain valid even though humanity will extend indefinitely
into the future. For an essay explaining the threat of nanotechnology, which
could include the means for achieving individual immortality, see the article
by Bill Joy at http://www.wired.com/ wired/archive/8.04/joy.html
Appendix D contains a description
of equations presented for those wishing to reproduce some of the preceding
material concerning population versus date matters.
─────────────────────────────────
CHAPTER 21
─────────────────────────────────
THE
PULL OF TRIBALISM THREATENS CIVILIZATION
Man is at bottom
a dreadful wild animal. We know this wild animal only in the tamed state
called civilization… Schopenhauer, 1851, Parerga and Paralipomena
(i.e., Aphorisms).
Introduction
In the previous
chapter I estimated that there's a 50% chance that humanity, as we know it,
will cease to exist sometime between 2100 and 2600 AD. In earlier chapters
I have tried to identify factors that may contribute to the disintegration
of civilization. In this chapter I will try to imagine how one scenario for
an unfolding disintegration may occur, and then present a theoretical unification
that should embrace all scenarios of this type. The specific scenario is
based on the notion that communists and fascists, with their primitive longing
for opposite aspects of the tribal lifestyle, are twin enemies of artisan‑created
civilization.
The closer we approach
the final population crash during the next few centuries the more accurate
will be our speculations about it. It is inevitable that future generations
will view speculations from this era as "outdated" ‑ and I acknowledge that
the following hypothetical scenario may never happen. But it at least contains
elements which have a credible claim for relevance.
Perhaps the biggest
disclaimer for the many endeavors in this book to discern civilization crash
scenarios, and an exploration of new directions for human evolution, is that
the world's population may not crash, and that to rely upon the Doomsday
Argument is to be misguided since it may not apply to the human population
situation. Therefore, the preconditions for the evolution of a new species
of humans who achieve liberation from their genes may therefore never exist,
and consequently there is a greatly reduced probability that a new species
of gene‑liberated humans could evolve. These are uncertainties we must live
with.
Scenarios Overview
The population of
Europe began to collapse
at about 200 AD, coinciding with the disintegration of the Roman Empire. Rome’s population fell
by a factor 10 during the centuries following 200 AD. Civilization is needed
to sustain a large population, and since civilizations are more quickly destroyed
than created, population collapses will be faster than their rises. The
important point here is that for the one well‑documented example of the world's
intellectual vigor and its population, the latter has a peak centuries after the former. For the reader's
convenience, I repeat two figures from the previous chapter that illustrate
this concept.
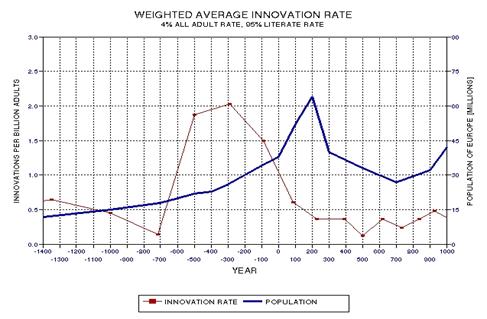
Figure 21.01. Repeat of Fig.
20.16, for a 1400‑year period centered on the Golden
Era of Greece (Chapter 20 has
a fuller explanation).
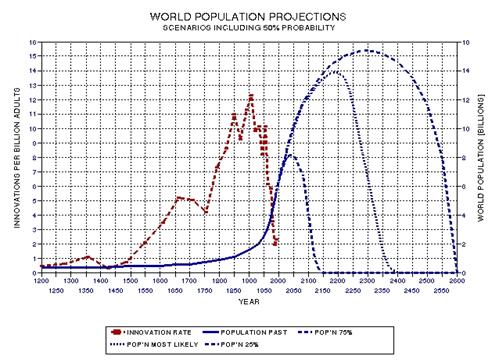
Figure 21.02. Another 1400-year
period, showing three hypothetical world population scenarios (Chapter
20 has a fuller description).
Innovation rate
and population during the current, post‑medieval period appear to be following
a pattern similar to the one associated with Classical Greece and the Roman Empire. In both cases
the population continues to rise after a peak in innovation rate.
The reader may find
it difficult to believe the precipitous drop of the innovation rate trace
in Fig. 21.02, but recall that the trace is a "per capita innovation rate,"
and is therefore influenced by the rising world population. In absolute terms
the rate of innovations is still increasing, but the world population is
increasing at a stupendous rate which causes the per capita innovation rate
is to drop.
I claim we might
gain insight into our future by studying the fate of the Golden Era of Greece,
which prepared the path to the Roman Empire (which ran its
course and was replaced by the Medieval Ages). At the height of the Roman Empire the influence of
Rome spread across the
entirety of that part of the world known to the Romans. After the "decline
and fall" the influence of Rome was greatly reduced,
and regions went their separate ways, like wanderers lost in a forest.
In each distant region the rule of Roman law was replaced by the rule of
local lawlessness. Grievances could again be settled by the action of small,
local coalitions, probably family‑based in most cases. In some ways this
resembled the state of affairs before the Roman Empire, when tribes had
two sets of rules: those that applied to others within the tribe, and those
that applied to outsiders (Spencer’s amity/enmity duality of morality).
I shall assume that
whenever the rule of law disintegrates, a population collapse begins. Consider
the 50% population scenario of Fig. 21.02, which has a population peak at
about 2200 AD. This scenario corresponds to a disintegration of the rule
of law throughout the world at that time. During the subsequent 200 years
the world's population shows a collapse to small values, as happened after
the Roman Empire's "decline and
fall." In this chapter I will adopt this specific timetable for describing
scenarios of how today’s global civilization might collapse.
Some of Civilization’s
Discontents
Let us "step back"
for a moment to review the long march of our human ancestors toward the glorious
civilizations of the Holocene Epoch. Hopefully this will help us discern
"big picture forces" which create and destroy civilizations.
Humans are presumed
to have lived in tribes during the past 1.8 million years of the Pleistocene
Epoch, dominated by glacial periods interrupted at 100,000 year intervals
by 10,000‑year interglacial warmings. The evolution of larger brains, occurring
at 1.8 million and 0.3 million years ago, is probably climate‑driven, ultimately,
but also associated with tribal adaptations to the challenge of living with
a variable climate. It was not the individual who faced a harsh environment,
but individuals living in tribes who faced harsh and variable environments.
The human mind must have evolved adaptations that allowed it to meet the
challenges of tribal living when climate became variable, causing landscapes
and hunting styles to change profoundly.
During the Holocene
interglacial warming, starting 11,400 years ago, those individuals prospered
who changed their connection to their tribe by adopting farming and herding
lifestyles. Gradually, a sedentary village way of living replaced the hunting
and gathering lifestyle. As weakened tribes coalesced into super-tribes,
then villages, cities and eventually the metropolis, civilization was created.
Civilized cities act like a magnet attracting rural dwellers to the glitter
of the city.
But life in cities
is fundamentally different from life on farms, just as life on farms was
fundamentally different from life in hunter‑gatherer tribes. Both transitions
brought the average person into contact with strangers on a daily basis. When
the farmer returned from his fields to his village home, or when he sold
his produce at a farmer's market, he was surrounded by strange faces. The
strain of seeing strange faces on a daily basis may have been even worse for
permanent city dwellers.
Recall that in the
tribal setting a person is surrounded by friendly, fellow tribesmen on a
daily basis, and the only time strange faces are encountered was during conflicts
with neighboring tribes. During the 1.8 million years of Pleistocene living
our ancestors must have evolved an automatic association "strange face means
enemy." Mental modules (i.e., neural networks) were created by the genes
to make this association, and those gene-created modules did not disappear
as soon as tribal living was abandoned. It is possible that even today we
become uneasy after encountering many strangers on a daily basis. We must
suppress these ancient instincts as we pretend to not dislike strangers. Yet,
we still have brain modules adapted to the ancient reality that "strangers"
mean "danger." A part of us must wish for a return to the tribal setting,
where we could count on a peaceful social setting, and where the need for
tribal cohesion produced a "brotherhood" of loyal social relationships.
It is important
to appreciate the abruptness of our transition away from tribal living. Compare
the 1.8 million years of Pleistocene living, or even the 4 million years
of human evolution following the separation from our chimpanzee‑like ancestors,
with the shortness of the Holocene's 11,000 years. Our
experiment with non‑tribal living represents less than 0.5% of the "human"
evolutionary experience. How much could our genome have changed during
the past 11,000 years? The genes coding for skin pigment can change during
times this short, and some aspects of anatomy can change (witness the native
American Indian, who left Asia at the beginning of the Holocene), but the
bulk of our human genome is identical to the one that was adapted to the
previous 1.8 million years of tribal living. We are forced to consider that
there are parts of our brains better adapted to tribal living than to life
in a modern civilization, and that in some ways people would feel more comfortable
living in a tribal setting than the modern civilized one!
What a poignant
realization! Civilized living is unnatural, and parts of ourselves wish to
return to tribal living, as if we still primed to hear that “call of the
wild.”
The implications
of this speculation are profound. Not only does it help us understand the
aimless frustration with life in these modern times, but it might help us
understand the political motivations of various "social movements."
Alternation Between Inter-Tribal Conflicts and Intra-Tribal Harmony
In order to understand
the subconscious feelings after being abruptly forced into a civilized world,
let us dwell upon the "feel" of tribal living. It will be useful to think
of tribal life as consisting of two flavors: things you do during intertribal
conflicts, and the things you do during the longer interludes between conflicts.
Conflicts can take the form of defending against attacks by neighboring tribes,
or they be the initiating of conflict by the
home tribe. All-out tribal conflicts may be rare, separated by years. The
more frequent conflicts can be described as “raids,” which are brief encounters
by small bands from a neighboring tribe that attack when they feel they
outnumber their victims and can overpower them with small risk to themselves.
These will usually occur near the periphery of the territory claimed by
a tribe, near contested borders.
The existence of
defensive palisade structures, dating to at least 30,000 years ago, are testimony
to the importance of the less frequent, large‑scale conflicts (Keeley, 1996).
For these inter‑tribal wars tribes would have relied on their best warriors
more than during opportunistic raids. The same warriors not only defended
the home tribe, but they are the ones who initiated tribal conflicts. Let
us appreciate the importance of the ancient warrior niche in every tribe.
I will assume that there is a "warrior" mentality, as well as an instinctive
appreciation of the warrior as a valued member of one's own tribe.
The majority of
tribal life consisted of peaceful endeavors, of activities that sustained
the tribe by providing food, shelter, tools, weapons and other economic necessities.
The tribe's survival hinged as much upon economic strength as on its war‑waging
prowess. Indeed, the ability to defend the tribe and wage war upon one's
neighboring tribe is determined by the productive payoffs of peaceful economic
activities. I will therefore assume that there is a peaceful producer mentality,
as well as an instinctive welcome of the producer members of one's own tribe.
As I have argued
earlier, late in the Pleistocene the artisan became one of the tribe's most
valuable assets. He was the full‑time toolmaker and weapon maker. For the
advanced tribes there was a place for the artisan who specialized in constructing
dwellings, storing food, gardening, and domesticating the occasional animal
that could be domesticated (goats, dogs, etc). Initially, tribes had few
full‑time artisans. When the climate warmed during the Holocene artisans became
more valuable, as they could more profitably pursue farming, herding and
other specialized economic activities. Therefore, I will assume that there
is an "artisan" mentality, as well as an instinctive (though grudging) welcome
of the artisan as a member of one's own tribe.
Three Mentalities
Enter Civilization
This was the condition
of tribal living that existed before the artisan created "civilization."
I have identified three mentalities that were important in the tribal setting
of the Late Pleistocene; there are others, but for now I want to focus upon
this most important triad. These three mentalities can be referred to as
1) the warrior, 2) the everyday producer, and 3) the specialist artisan. The
distinction between the "everyday producer" and the artisan, both of whom
are producers, is that the first has no unique skills and is pressed into
battle during tribal conflicts, whereas the artisan is "protected" during
tribal conflicts. Because of a resentment over
this the acceptance of the few artisans in each tribe was made grudgingly.
Now, consider how
these three types of people would react to the spectacular explosion of civilization.
Since "civilization" is the handiwork of the artisan, he would feel comfortable
in this new setting. However, the warrior and everyday producer would feel
somewhat “left out” and “out of place.” They would feel uneasy for being
forced to endure daily encounters with strangers, since strange faces "trigger"
subconscious anxieties. The loss of tribal rituals and a world of surrounding
spirits would also contribute to a feeling of dissatisfaction with civilized
living. Within the minds of people who are inclined to the warrior and everyday
producer life, there would be an ambivalent feeling about the new civilized
state. A subconscious "homesickness" for the tribe may become stronger the
more civilization advances.
But the homesickness
for tribal life would be slightly different for the warrior type and the
everyday producer. The warrior would miss his crucial and celebrated role
in defending his tribe and attacking others. The everyday producer, on the
other hand, would miss the ambience of a peaceful interlude of cooperative
comradeship. These best of times were peaceful, friendly, cooperative, and
infused with an atmosphere of loyal togetherness that sustains effective group
living. At these times it is fair to characterize one's fellow man as "peaceful"
or "noble."
Birth of Communism
There's a "place
in the brain" for the concept "noble savage." Jean Jacques Rousseau felt
it, as do all present‑day Marxists and communists. This faded memory is easily
resurrected, and Marx played on this with his interpretations of history.
He wrote that the “class struggles” characterized modern capitalist societies,
and this struggle was due to the corrupting influence of culture. Moreover,
since he believed that human nature remained noble, a change of culture could
return us to that dreamed of state where everyone worked harmoniously together
for the good of the group. His wish to return to commune living, and his
belief in its feasibility, are manifest expressions of a subconscious nostalgia
for our million-year experience with tribal living. I believe his perspective
can be distilled to the following: “If only people could break the shackles
of a dysfunctional culture (i.e, civilization), then all
Mankind could return to simply living together in ancestral peace and harmony.”
Birth of Fascism
Marx overlooked
the other half of human nature, namely the human appetite for inter‑tribal
conflict. Those who most strongly longed for intertribal conflict are today’s
fascists! The fascist's gripe with civilization is that it is too peaceful,
with too much civil behavior, codified by artisan‑written laws and enforced
by police and courts, which deprive the fascists their most important tribal
role. Whereas no one celebrates the criminal today, everyone celebrates the
warrior ‑ even though both thrive on mayhem and murder. The fascist thus
wishes for a breakdown of society, and a return to the kind of anarchy that
so pleased his ancestors.
The fascist also
dislikes civilization’s celebration of individualism. Fascists instead embraced
“authoritarian order, based upon the subordination of the individual…” (Encyclopaedia Britannica, 1977, 7, 183). They also have “a definite attitude of mind
that exalted the fighting spirit, military discipline, ruthlessness and action…”
(op cit). “The
Italian slogan [under dictator Mussolini] to
‘believe, to obey, to combat’ was fascism’s antithesis to ‘liberty, equality,
fraternity’” (op cit). Of course, “liberty, equality, fraternity”
grew out of the French Enlightenment, which in turn was an expression of
the artisan’s value system.
Chapter 19 defined
fascism using a statement uttered by Benito Mussolini, who urged a combining
of business and state leaders for authoritarian rule of a nation so that
it may pursue “belligerent nationalism.” The part about combining business
and state leadership is simply a means for hijacking a nation from a democratic
embrace so that it may be set on the belligerent nationalism course. “Belligerent
nationalism” really refers to the primitive drive for initiating inter-tribal
warfare for the goal of enlarging a tribe’s dominion over a larger territory
and subsuming rival tribes. A modern name for this desired en-point is “empire.”
In this chapter I will emphasize the warrior aspect of fascism since it springs
from the deepest parts of a person’s instinctual inheritance.
Communists and Fascists
as Twin Enemies of Civilization
The fascist and
the communist have something in common, even though they are traditionally
assigned opposite ends of the political spectrum. Both wish for the destruction
of the civilized state and a return of society to a tribal lifestyle. Both
harbor a deep hatred of the artisan, whose creation they wish to destroy.
The early part of the 20th Century was wracked by the simultaneous
efforts of fascists and communists to destroy the society that had thwarted
the expression of one's natural self, and denied realization of a half‑remembered
idyllic past where these people were important.
The forces of discontent
unleashed during the early 20th Century, between World Wars I
and II, caused many thoughtful people to worry about the end of civilization.
Fortunately, these destructive forces were subdued and returned to subconsciousness
after World War II. But they remain as an unnoticed wish, partially hidden,
and these desires are a serious threat to civilization. I predict that the
21st Century will see a replay of the assault of tribalism upon
civilization!
If the Fascists
had Won World War II
If "civilization"
had not won the Second World War, which some suggest was a "close call,"
then we might now be witnessing a slide into a new Medieval Age. The shape
of this world would have begun with Hitler’s version of fascism, not Stalin’s
version of communism. Even Lenin acted like a fascist, and his embrace of
communism seems more a convenience than a fervently believed‑in ideology
(Server, 2000). The decline and fall of today's civilization is therefore
likely to resemble, in its early stages, the scenario one can imagine if victory
had gone to the three Axis fascisms: Nazi Germany, fascist Italy, and racist Japan. We may assume
that Germany would have subjugated
the vanquished, given their superior military; so imagine a world styled
upon the Nazi dream.
It would be an understatement
to say that Nazi Germany was racist. They were racist in the classic tradition
of tribalism. The first rule of the "tribal mentality" is to hate strangers
and love your fellow tribesmen. It is common in primitive languages to employ
a term for everyone belonging to other tribes that is dehumanizing, such
as "dog" or "snake" (Chagnon, 1992). These epithets help arouse the right
attitude for acting unmercifully toward the "others" during tribal conflicts.
During World War II, as in other times of war, the same technique was used
to help arouse the required tribal hatreds and brutal behaviors. The Nazi
victors would have continued their "ethnic cleansing," and it would have
become a global project. The Nazi killing of non‑Aryans might have gone on
for decades.
The “Nazi Dark Age”
would have run out of energy before world ethnic cleansing could be completed,
given the immense magnitude of the task of exterminating billions instead
of millions. I believe this would be the case because the fascist is motivated
to behave in ways that "made sense" when one's enemies were limited to the
troublesome neighboring tribe. Exterminating the rest of the world would
become a weary task, in which the confused Nazi would probably lose interest
eventually. After abandoning the ill‑conceived goal of world ethnic cleansing,
the Nazis probably would have settled for plundering the rest of the world
‑ which they had already started during the war.
So, imagine a world
in which non‑Aryan populations lay decimated, in which world commerce has
been converted to a one‑way transport of plundered goods, and in which artisan
initiative is no longer rewarded as it is in civilized societies. The Nazi
Dark Age would have degenerated from organized racial exterminations, to
plunder, to chaos and eventually disinterest. The rest of the world would
have been robbed of the resources and incentives to reconstitute itself as
a civilization.
This, I submit,
could still happen. Even at a time when we know that the hidden hand within
the glove of tribal cruelty is the genes. The genes “act” as if they cannot
tolerate competing alleles, and this intolerance
is what fashions all animal behavior, including human animals. The greater
insight that we now have concerning human behavior will be a puny force in
the face of the juggernaut of evil primitive forces. No matter how much the
artisan learns, no matter how powerful his insights in the coming decades,
he is destined to be squashed under fascist boots. These boots have a blind
wish to march, with the innocence of a noble savage heart, to a societal
state which a vague memory clings to, of the way things were, and should be
again: the beloved tribe to which all but the artisan
mind is well adapted.
The shape of the
new Armageddon will be qualitatively different from any alternative unfolding
of the 20th Century. For in this 21st Century the world's
population is 3 times larger, global oil production “peaked” during the
21st Century’s first decade, natural resources are nearly exhausted
and require more technology and energy to extract, the environment is more
contaminated than at any previous time, small terrorist groups are a greater
threat than a complacent First World has the resolve to reckon with, there
are larger pools of diseases ready to hitch a ride with people moving globally
on a daily basis, and on and on.
The new "world system"
has many more variables than at any previous time, and I feel overwhelmed
by the daunting task of predicting in any detail how the next Dark Age will
unfold. All I feel certain about, though, is that our primitive ways of thinking,
that served our genes well for millions of years of tribal living, will
provide the energy for the micro‑motives that together will produce macro‑behaviors
longing for the tribal setting. And since our primitive desires will never
be in alignment with civilized living, civilizations will always be transient
affairs.
The Chinese Empire
Although I hesitate
to make specific predictions, I will present one possible scenario.
As I write this
(mid-2008) China’s economy is booming
and America’s is sputtering!
China is investing in
a military build-up that seems disproportionate to present threats. An army
is created for one purpose (killing people) and if it achieves sufficient
strength to fear no foe, and if it is unused for a long time, political forces
stemming from that army will “rattle its sabers” and give courage to ambitious
business leaders and politicians to instigate an international crisis with
high stakes potential benefits. China appears to be “itching
for a fight” to take over Taiwan, which they have
steadfastly claimed is a part of China and not a separate
country. If China invaded Taiwan (let’s say in 2020)
it would be logical for there to be no response from America. This is because
the American economy would collapse almost immediately if China retaliated by demanding
payment of its U.S. Treasury bonds, or even stopped buying new ones.
It may be logical
for America to refrain from defending Taiwan but internal American politics
will not allow a non-response; it would be too humiliating for whoever is
president of America (especially if it’s John McCain). So imagine that America attempts to defend
Taiwan, and China responds by stopping
the purchase of U.S. Treasury bonds, demands payment for the U.S. Treasury
bonds that it holds, manipulates its currency to make American imports
of Chinese products more expensive, and does other things to punish America. The American economy
would collapse, China and other foreign
countries would buy American assets, such as mines, ports, financial institutions,
and little by little America would devolve into
a series of foreign-owned colonies.
How ironic for America
to start out as 13 British colonies, and end up as a dozen or so colonies
controlled by China, Japan, India, Saudi Arabia and other Middle Eastern,
oil-rich countries. The American colonies would provide resources for use
in the home countries. China needs copper, cement,
lumber, other raw materials, and food, such as grain, corn and cattle. American
mines would be owned and run by Chinese companies, America’s forests would
be cut down by lumber companies owned by China, and farms would
be run by China. America’s rail and highway
infrastructure would finally be upgraded in order to transport America’s resources to
ports where Chinese ships would be waiting.
Americans would
find employment with Chinese companies. The Chinese language would be taught
in schools, replacing Spanish as the most popular “foreign” language. Since
most Americans would be unemployed, some of them would be hired to provide
security for the new colonial commerce. Immigration that now is from Mexico to America would reverse
direction. Those who could not find work with Chinese or Saudi Arabian companies,
and who saw no future in emigrating to Mexico and were prevented
from emigrating to Canada, would simply kill
each other, starve and die.
At this stage in
China’s growth it would
be fair to refer to China as an empire. Every
empire has a finite lifetime. This is because the rulers of an empire are
the ones who had the fascist outlook. They rise to power because in a powerful
nation individual freedom is incompatible with social stability. China has a “head start”
in controlling individual freedom. This is partly due to a fundamental flaw
in the Chinese character, the willingness to passively accept feudalism,
a king’s rights and fascism – which are the same in their demands on the individual.
(Why else is there a scarcity of real scientists in China; a scientist must
have a strong individualistic personality. Engineers are another matter, they can be submissive and still be effective)
In this scenario
China evolves into a
fascist state, let’s say sometime in the middle of the 21st Century.
Far flung colonies throughout the world provide China with raw materials
and food. China has scores to settle
with Japan, Korea and Russia, so to suppress
growing civil unrest over a straining economy in a too-large empire China begins to wage
wars with its neighbors. At this point, consider the state of the world to
be similar to the state of the world during World War II, when Germany was threatening
world domination using their military supremacy. China wins all its wars,
and extends its empire to the entire world. Now, refer to the previous section’s
scenario and run it with the word “China” replacing “Nazi
Germany.”
This is obviously
a hypothetical scenario that is unlikely to unfold with the same details
I used to illustrate things that could happen. Other scenarios could be imagined
in which India become an empire instead of China. The world’s civilized
condition may revert to a Middle Age barbarity during the 21st
Century; or it might be the 22nd. The great descent into Dark
Age barbarity might occur in the year 2050, or 2200, or 2400 AD, but surely
the time will come when the civilization we know and pretend to cherish will
succumb to dark forces more ancient than any human life, and more compelling
than any noble human aspiration. The accomplishments of Beethoven, Schopenhauer,
Darwin, Bertrand Russell, E. O. Wilson, and all my other heroes, every noble
Beauty and Truth which they strove to bring to life, will fall into the ashes
of a dying civilization, perhaps never to return again.
This dark scenario
is but one of many that could be constructed from the discontents of those
forced to endure “civilization.” The "backward pull toward the primitive"
has many faces; and the communist/fascist variety is merely one example.
People who are less intelligent and articulate may prefer anarchy, the great
leveler that forces those with ability to suffer equally with those who lack
ability. There are religious fanatics who feel that their cozy emotional nest
is threatened by what the rest of us call ideas and progress. Each malcontent
will react to civilization differently, but they all will feel, at a subconscious
level, that there’s something wrong with civilized life, something is out
of place, and to fix things that went wrong during our historical journey
we must somehow return to an idyllic past condition, a Garden of Eden. And
this means dismantling civilization, as if by doing that we will thereby free
ourselves from alien, corrupting influences. Alas, that darned world that
artisans created will be destroyed.
Cultural "Reverse
Evolution"
This section presents
a "unifying theory" which endeavors to embrace each category of dissatisfaction
with civilization. It accounts for how most people are subconsciously unwilling
to leave the emotional comfort of the ancestral environment. Regardless of
civilization's material bounty, regardless of the plethora of awesome insights
which spring from civilized intellectuals, civilization nevertheless presents
an unwelcome mat for the human emotions. It is as if many people are dragged
into the civilized state who are silently screaming and digging in their
heels ‑ waiting for an opportunity to return, by whatever means.
In the Chapter 4
genetics tutorial I described something called "reverse evolution." The basic
idea for this new thought is that the most recent mutations in a genome are
the most vulnerable to loss, and that the evolutionary "advance" that they
produced is the most vulnerable to "evolutionary reversal." This is because
the most recently acquired genes have had less time to acquire "allies"
for genetic survival (apologies for putting it this way, which is quite anthropomorphic).
The longer a gene is "in place" in a genome, the more it becomes depended
upon by newer mutations at other chromosome loci. The new mutations provide
their contributions to fitness only for as long as certain other pre‑existing
genes remain present. If a pre‑existing gene is deleted, then not only does
the individual lose the trait it brought into existence, which was the original
justification for its selection, but the individual also loses traits that
later genes brought into existence when they depended upon the presence
of the first gene for its proper expression. This intertwining of genetic
effects is captured by three complimentary terms: Pleiotropy, polygenes
and entrenchment. "Pleiotropy" refers to the fact that a single gene can
influence many traits, "polygenes" refers to the fact that a specific trait
requires the presence of other genes,
and “entrenchment” describes the situation of old genes being difficult
to get rid of because they have more newer genes depending on their presence
for them to work properly than is the case for not-as-old genes.
Because an "entrenched"
gene has many "allies" for resisting meddling by hypothetical future mutations,
the entrenched gene has an advantage when a rival begins to compete with
it. Let us use the term "unentrenched" to refer to a recently acquired gene.
Unentrenched genes are unlikely to have allies, and are therefore more
vulnerable to "challenge" (I ask the reader to understand that my use of
anthropomorphic terminology is done for brevity, and any reader who is bothered
by this anthropomorphism must provide his own long‑winded reductionist version.of
the thought I mean to convey).
When environments
change, the unentrenched genes are the ones most likely to be replaced by
allelic challengers. This is especially true when the environment changes
back to an earlier one, to an environment that existed before the unentrenched
gene came into existence. Indeed, if the unentrenched gene came into existence
in response to a change from Setting A to Setting B, and if Setting A returns,
the unentrenched gene is a very likely candidate for being replaced with
the gene that it had replaced ‑ assuming some of them still exist in a few
individuals.
"Civilization" is
a product of a fast‑paced cultural evolution, made possible by a much slower‑paced
genetic evolution. The genes coding for a specialized left brain (LB) rely
upon a serial processing architecture of interacting, self‑contained neural
networks. The right brain (RB) relies upon a more primitive neural architecture
of parallel processing. LB logic led to the accidental creation of a social
arrangement we call "civilization." I shall assume that contemporary humans
are little more receptive to civilized living than those of our ancestors
who were present to usher in the Holocene. In other words, I shall assume
that "civilization" was created by an unfolding of an ever‑more complex culture
with few genetic accommodations.
Cultural evolution
is constrained by the genome; hence, the potentially fast-paced evolution
of culture is constrained by the slower-paced evolution of the genome. Genes
determine what elements of culture, called culturgens (Lumsden and Wilson,
1981), are theoretically possible, and probable. After a culturgen has been
universally adopted throughout a gene pool the social environment is changed,
and this change may create new opportunities for genes. The co‑evolution
of genes and culture is a young science, but some rudimentary properties of
it are already understood.
Since culturgen
frequencies can change much faster than gene frequencies, it is possible
to consider cultural evolution in the absence of genetic evolution. To my
knowledge, the matter of "cultural evolutionary reversals" has not been studied,
or even mentioned, in the literature. The term "reverse evolution" is recognized
as applying to genes. So let us use the term "cultural evolution" to refer
to culturgen changes that occur faster than gene frequency changes.
Chapter 4 described
the results of a genetic study of fruit flies by Teotonio and Rose (2000),
showing that they can achieve 90% "reverse evolution" (for many traits) in
only 10 generations! The "recovery lines" appear to be linear, so a 10 %
recovery can be accomplished in just one generation! These data are for "unentrenched"
gene alleles. Surely, the recovery times for unentrenched human culturgens
would be much shorter, as measured in generations. Adopting a human generation
time of 30 years (Pang et al, 1998), pure cultural evolution
could achieve a complete reversal in much less than 300 years, and partial
reversals in about 30 years.
In the context of
the rise and fall of civilizations, it is important to ask if pure cultural
evolution can account for the rise and fall of civilizations, or must genes
be invoked in a way that requires a mixture of cultural evolution and genetic
evolution to give an accounting of the rise and fall of civilizations. We
should be prepared for an answer such as "a mixture of genetic and cultural
evolution causes civilizations to rise, but almost pure cultural evolution
causes them to fall." This would be compatible with the fact that civilizations
rise with a timescale of approximately 1000 to 2000 years, whereas they fall
with a timescale of 200 years (i.e., 30 to 60 generations,
and 6 generations).
Entrenchment of
Culturgens: Archosis, Taboos and Conformism
Is there anything
analogous in cultural evolution to pleiotropy and polygenes? Can it be said
that one culturgen affects others? Or that many culturgens contribute, or
are required by, one cultural trait. What we're after is an answer to the
question: Do culturgens interact in such a way that recently acquired culturgens
are easier to shed than longstanding ones?
I think the answer
to all these questions is "yes." Consider, for example, the adoption of fire
for preparing food. With just this one culturgen many new lifestyles became
feasible. Foods that were not digestible become edible; foods that spoiled
easily can be eaten for a longer time; campfires keep dangerous animals
at a safe distance, allowing for larger groups of people to live in areas
where large predators also live, etc. Clearly, the adoption of this one
particular culturgen changed the feasibility of adopting many other culturgens.
This one example shows that not only is there a culturgen counterpart for
pleiotropy, but one culturgen can affect the fate of other culturgens.
The longer the fire
culturgen is used, the more other culturgens will be adopted that depend upon
the existence of the fire culturgen. Therefore, for any tribe whose culture
has relied upon the use of fire for a long time, over time there will be
greater penalties for abandoning the fire culturgen. It is therefore inescapable
that there should be a tendency for culturgens to exhibit the same property
of genetic evolution reversibility, that the longer a culturgen remains adopted
by a society the more “entrenched” it becomes over time. Indeed, since this
principle can be expected to apply generally, regardless of the specifics
of the culturgen, there may have been rewards for our ancestors to adopt
those culturgens which punish thoughts and acts that appear to defy ancient
culturgens. Rituals and traditions may thus be revered merely for their longevity,
and the concept of "taboo" may itself become a valuable culturgen.
As an aside, let
us acknowledge the seminal work of Weston LaBarre, who introduced the term
"group archosis" (LaBarre, 1984) to refer to "nonsense and misinformation
so ancient and pervasive as to be seemingly inextricable
from our thinking. ...A frightening proportion of all culture is arguably
archosis, more especially sacred culture." Dan Sperber, taking a similar
position (1985), went on to suggest that genetic evolution produces mental
dispositions with "side effects" that have marginal adaptive value, with religion
being one example (as recounted by Edgerton, in Sick Societies,
1992, pg. 53).
“Conformism” was
described in Chapter 10. Culturgen entrenchment could be the theoretical
explanation for conformism.
If these speculations
are true, then it will also be true that recently acquired culturgens will
be un‑entrenched, and they will be the ones most easily discarded. This is
the feared position I have been working toward in this section. For, to
the extent that civilizations are creations based upon the adoption of new
culturgens, the robustness of a civilization, and its ability to rebuff cultural
assaults, will be weak. The genes have created a brain that is demonstrably
capable of creating a civilization; but since the genes have no experience
with sustaining civilizations there can be no guarantee, and indeed there
should be little assurance, that the civilization will endure the continual
challenge of people who are born with a nature that feels more comfortable
in the primitive, tribal environment. The amazing thing about civilizations
is not that they seem to always collapse, but that they ever are created
in the first place.
Role of Time in
the Rise and Fall of Civilizations
Figure 20.01 shows
that the ancient Greek civilization may have "peaked" at approximately 300
BC, and the Greco-Roman civilization was in collapse starting at ~200 AD
‑ when, ironically, the region's population had reached its peak. Using "per
capita innovation rate" as the measure for the strength of their civilization,
it had lost 90% of its peak strength in 400 years, which corresponds to about
14 generations. (If Greek civilization had not been adopted by the Romans,
the loss might have occurred closer to 10 generations). Therefore, we cannot
rule out the possibility that gene frequency changes did not occur, and
contribute to the decline. The decline may in fact have been a mixture of
genetic and cultural reverse evolution.
The "rise" of Greek
civilization appears rapid in the figure. In 200 years the "per capita innovation
rate" rose from a background level to the peak. I suspect that "per capita
innovation rate" is inadequate for revealing the role of underlying genetic
changes that eventually led to the apparent abrupt blossoming of civilization.
As Carl Sagan pointed out (1980, p. 174), a region called Ionia was a hotbed for
new ideas and inventions starting in the 6th Century BC; these
practical-minded thinkers introduced the rudiments of the philosophy of
reductionism. We can only speculate that perhaps the Ionian population was
based upon a diaspora of Minoans, who had to flee their island homelands
when the volcanic eruption of Thera destroyed much of the infrastructure
of their civilization in 1628 BC (Pellegrino, 1991) and left them helplessly
exposed to the invasion of Mycenaeans from the mainland. If the Ionian and
Greek civilization is a continuation of the Minoan civilization, then there
is a case for a 3000‑year rise phase. Such a long time, representing 100
generations, could easily have been influenced by gene frequency changes.
We are left with
an ambiguity on how to partition credit for the rise and fall of civilizations
between the two candidate root causes, genes and culture. My personal preference
is to credit the "rise phase" to a co‑evolution of genes and culture, and
to blame the "fall phase" almost exclusively on culture. During the rise,
new culturgens improve the congeniality of civilization‑promoting genes,
which lead to more civilization‑promoting genes, which in turn leads to a
greater congeniality of the social environment to culture‑promoting genes,
and so forth. This process may sometimes lead to the creation of a civilization.
At some point during
this process, a culturgen crucial to the well‑being of civilization is challenged
and discredited. This fateful challenge may not make sense to a thinking
brain, but most people within a culture are unthinking. An old way of believing
and behaving simply seems more attractive to enough of the populace that the
discrediting of the new ways is accomplished, almost effortlessly. The loss
of a new, un‑entrenched culturgen is a case of "reverse cultural evolution,"
which has its counterpart in "reverse genetic evolution." The details of
what must be a large number of "reverse cultural evolution" episodes is open
to speculation, but when a critical loss occurs the fate of the targeted civilization
is sealed, and the unraveling proceeds as if destined by compelling but
unseen forces. It may be that the new culturgens that created the civilization
had been resisted all along, but only after the populace acquired power did
their opinions matter and produce destructive effects.
Sigmund Freud wrote
Civilization and its Discontents (1930) at a time when there
was a gathering storm in Europe that caused many
intellectuals to become concerned over the prospect that Western Civilization
might not survive the mischief of a flawed human nature. The climate of opinion
favored serious questioning of the inevitability of civilization as an enduring
state for humans. Freud cited the human heritage of animal instincts from a tribal setting,
and he questioned whether civilizing forces were capable of controlling the
impulse for uncivilized behaviors. One famous line from the book is “…men
are not gentle, friendly creatures wishing for love, who simply defend themselves
if they are attacked, but that a powerful measure of desire for aggression
has to be reckoned as part of their instinctual endowment.” Incidentally,
Freud would probably be receptive to the sociobiological paradigm, for he
wrote “…a psycho-analytic, that is, genetic explanation…” (op cit., pg. 10).
Chapter Conclusion
Whatever insights
are eventually revealed by further study, at this time we can say with some
confidence that the pull of primitive ways of thinking and feeling produce
a powerful and subconscious longing for those well‑established culturgens
that evolved in a tribal setting. Further, we can say that this "emotional
longing for the past" drive reversals in cultural evolution that eventually
destroys each civilization. Since going backwards to the familiar is emotionally
more inviting than sustaining an uncomfortable present, or going forward
into an uncertain future, the primitive parts of men's minds will always try
to unravel the civilization woven by the creative efforts of a few brave men.
The unraveling is driven by RB-styled people, who resent the emotional discomforts
of the civilization created by LB-styled artisans.
I suggest that the
rise of a civilization is due to a favorable co-evolution of genes and culture,
whereas the decline is due only to the loss of recently acquired culturgens.
If what goes up
eventually comes down, then the "gravity" of human affairs is the wish for
emotional comfort, the feeling of a primitive pull to a vaguely‑remembered
tribal setting. In short, it is the desire to answer a profoundly human "call
of the wild!" And if this requires destroying whatever stands in the way
of the return, which happens to be civilization, then "so be it."
─────────────────────────────────
CHAPTER 22
─────────────────────────────────
LIVING WISELY ‑
SEEKING POSITIVES
"It is in human
nature to think wisely and act in an absurd fashion."
Anatole France
"Nature, Mr. Alnutt,
is what we were put here to rise above." spoken by Katherine
Hepburn to Humphrey Bogart in the movie The African Queen.
Human Nature was
formed to benefit our genes in an ancestral environment, and it has always
been a bad guide for the individual; it is an especially bad guide for individuals
in the modern environment.
The boxed statement
is this book's guiding thought for the person who wants to fine‑tune their
personal philosophy for living.
It is important
to have this thought in the background of our thinking as we consider the
merits of the everyday push and pull of our emotions. The enlightened person's
goal should be to understand the "ultimate causation" explanation for the
origin of thoughts and emotions. This challenging task can be done best in
calm moments. The process should also be the basis for the amendment of
one’s "value system."
Mount Cognoscenti
The more one understands
the world, the greater the challenge for reconciling internally‑generated
drives and goals with insight and logic. The person climbing "Mount Cognoscenti" at some point
arrives at an altitude where he begins to see the "existential dilemma."
The dilemma starts with an acknowledgement that life is inherently without
purpose, after eschewing the plethora of false goals meant only to serve
our genes. "Salvation" takes on a new meaning. How does one re‑invent a life?
How can an individual who has discredited gene‑contaminated goals invent
new ones which make sense and have value to a mind disciplined by logic? If
all values are ultimately derived from tricky genes, how can an individual
invent new values that are free of genetic trickery? As Nietzsche argued,
when God is at last dead for Man, when he feels surrounded by the darkness
of an uncaring universe, how does one go about inventing new values to live
by? The dismay following these questions is an "existential dilemma."
Throughout history
individuals have recorded what now can be seen as groping attempts to confront
the existential dilemma. The dilemma can only be sensed by those who are
able to see themselves as "individuals" ‑ entities with distinct identities,
theoretically separate from the crowd. Early societies had cultures drenched
with mysticism, and naturally the first groping attempts to understand the
individual's existential dilemma were greatly influenced by mystical thought.
It is an awesome irony that there are superficial resemblances between the
wisdom of early mystics and the new scientifically‑based wisdom that I espouse.

Figure 22.1 The two paths
to wisdom sometimes converge.
It can be disconcerting
when someone else arrives at the same destination after traveling a less
disciplined and less arduous path. The above cartoon (by my high school friend
Frederick L. M. Doll, 1999) suggests that a left‑brained scientist’s insights
resemble those of a right‑brain mystic. For example, the physicist's F=ma conception of a mechanical universe states
that every particle in the universe is connected to every other particle by
the four forces of physics (gravity, electromagnetism, weak and nuclear).
In other words, the universe is a fully-interconnected system in which everything
affects everything else, and in every corner of the universe things unfold
in response to the influence of every other thing in every other corner of
the universe. This description has an amazing resemblance to a tenet of Eastern
Thought, that everything that happens in the world does so in accordance with
a universally interconnected "flow."
For another example,
consider the longstanding quest for salvation. I remember when I was with
a high school friend (the same person who drew the cartoon, above) and we
encountered the phrase "What shall we do to be saved?" in Pilgrim's
Progress (page 64, a significant number in math, as I noted at the
time). We laughed mockingly at the quaint question, realizing that it referred
to salvation from a non‑existent Hell, yet the same question can be recast
with a modern sociobiological and existential context, which I am in effect
doing with this book.
Life'a a Funny Proposition,
Indeed
As the George M.
Cohan song says, "Life’s a funny proposition, indeed!" We're not "supposed"
to understand why we exist, for to do so would put one on the road to emancipation!
Our minds are an assembly of modules for getting a job done, which is to
promote genetic longevity, sometimes at the expense of individual welfare.
There can be no coherent understanding by the character at the center of this
predicament until he strays into the forbidden territory of logical, left‑brain
insight, and is lucky enough to discover the secret genetic tricks that have
victimized him. It is no accident that the biggest questions surrounding the
meaning of life appear to be inexplicable for most people. Each person must
"know thyself" in ways more profound than Socrates could have imagined.
Life is Backwards
Life is "backwards"
in the sense that only near the end of life do we have some sense of how
to approach life wisely. It is ludicrous that cruise ships are filled with
old people, who have the money but are losing the energy to explore the world.
If "youth is wasted on the young" then wealth is wasted on the old. Health
and wealth are too often found at opposite ends of life.
Risk Aversion is
Backwards
Another oddity of
human nature can be found in a pattern of "risk aversion." The young, who
have a whole life in front of them and therefore have the most to lose from
mistakes, seem prone to taking risks with an abandon that ignores large potential
consequences. The caution they should practice can only be found in the old,
who have the least amount of life at risk. Perhaps as people age some of
them begin to break loose from the genetic grip and realize that risk-taking
is for fools.
The Genes are Never Satisfied
As Schopenhauer
wrote, “…how insatiable a creature is man! Every
satisfaction he attains lays the seeds of some new desire, so there is no
end to the wishes of each individual.” “For intellect
is fundamentally a hard-working factory-hand, whom his demanding master,
the will [genes], keeps busy from morn to night.” (Schopenhauer, 1851, pg. 127). We serve the genes, and
no matter how much we’ve accomplished they are never satisfied. Schopenhauer
bemoaned man’s preoccupation with attaining what is close enough to touch
but not close enough to grasp, instead of appreciating that greater wealth
of what is already possible. When I retired I placed a sign in my office “Go
thy way, eat thy bread with joy, and drink thy wine with a merry heart, your
work is done.” (Ecclesiastes, 9:7, altered)
Wellness Begets
Wellness
Another of life's
little ironies is that the better off one is,
the better off one becomes. The converse is also true
– the worse off someone is, the harder it is to recover.
Over the years,
after noticing many life paths, I have come to realize that the world is
a great place ‑ provided you are physically and emotionally healthy. It takes
health to stay healthy, just as it takes wisdom to gain wisdom. To know that
exercise promotes health is only useful to the person who is healthy enough
to exercise. The healthier a person is, the more
feasible and fun it is to exercise, and the more that exercise improves
the person's health. Similarly, the smarter the person the more able he
is to learn and become smarter from such places as the internet and judicious
reading. But the person who never learned how to read, for example, has
lost many opportunities for new learning. Thus, there is a tendency for people's
mental and physical health to continue to improve if it starts out above
average, or to degrade if it starts out below average. Over time, the rich
get richer, and the poor get poorer ‑ in more respects than monetarily. There
is something inherently "unfair" about these life destinies; most people sense
this but can't express it. This version of how the world works is captured
by the following motto: "Unto every one that hath shall be given, and he shall
have abundance: but from him that hath not shall be taken away even that
which he hath."
The More One Knows,
the Less There is to be Known
The more a person
lives life with eyes open, the less of it that makes sense. To cite an oft‑quoted
commentary on the nature of the cosmos by a Nobel prize‑winner, "The more
comprehensible the universe becomes, the more pointless it seems." (Steven Weinberg, 1977). To
follow the crowd, or even to follow one's predispositions, is the quickest
path to folly. There is wisdom in the perverse method of searching for
Truth which states that you start with beliefs sustained by the masses,
and turn them upside down. The mystics sensed the fraudulence of common
beliefs, and fled to the quietude and clean air of the mountains. I share
this instinct, but do so guided by a left brain instead of a right.
Don’t Follow the
Crowd
Don't follow the
crowd! That should be one of the first rules for living. The crowd does what
the genes want done. At least, this is the case in environments resembling
the ancestral one. And since some of our genes are outlaws, anyone following
the crowd unthinkingly is open to fraudulent exploitation by these tiny "outlaws."
It is especially
difficult to instill this idea in children. They take their cues from peers,
while ignoring good parental advice. Their desperation to "belong" is so
strong that even an enlightened parent can feel helpless. Thankfully, most
of us survive the silliness of youth, and get second chances to face foolish
fashions. By adulthood there are no excuses for failing to be true to one’s self. As said by someone I overheard in a bookstore,
"When you live by someone else's standards, you betray your own."
Be Careful What
You Wish For
In retrospect, I
am thankful that in childhood the Forces of Fate denied me some cherished
longings. A part of me, a dumb part, wanted to be popular with the prettiest
girls. Thankfully, this was not to be, and my interests turned to less popular
aspirations ‑ such as making telescopes and home‑made cameras for photographing
the planets, auditing university courses while in high school, and taking
hitch‑hiking camping trips every summer. Only later did I recognize the wisdom
in the saying "Be careful what you wish for, for you may get it." There
must be a part of me that does battle with residual primitive wishes, for
it counsels that "When you get what you want, you deserve what you get."
Sayings like these help discredit what is bad for us, that comes from within
us.
Let the World Leave
You Alone
Someone on the street
asked “Are you Clint Eastwood?” and Eastwood replied “Only when I’m alone!”
The greatest gift to an individual is to be left alone, even if it means
feeling unwelcome in a dumb world of hand-me-down beliefs and values. A resourceful
individual will invent new values to live by, and if he is lucky they will
be better. As Nietzsche counseled, start worrying when people claim to understand
and agree with you. You may think that Life dealt Nietzsche lemons, so he
made lemon aide; if so, it was a lucky deal.
Beware Inner Wisdom
Ironically, whereas
my first specific advice is "Don't follow the crowd" I am also counseling
"Don't follow your inner self." Neither the crowd nor one's inner self can
be trusted, since both are creations of the genes - some of which are outlaws.
So what's left if
you shouldn't follow either the crowd or your inner self? My advice is "Don't
follow anything; start thinking for yourself!" In other words, start relying
upon left‑brain insights, and figure things out for yourself. Following is
for fools; wise men blaze their own trails.
Aesthetics
I will accept the
charge of being an aesthete! Aesthetics is as old as the record of human
thought. From the ancient Greek philosophers to 20th Century philosophers,
and especially during troubled times, there has been a place in the mind where
this theme resonates. Irwin Edman’s The Contemporary and His
Soul (1932) has given eloquent expression to this haven; it is an eternal
“place” that an individual can create when surrounded by tumult and confusion
(see Chapter 24 for more on aestheticism).
Brief Encounters
The book Brief Encounters by Coleman and Edwards (1980) states what should
be obvious to every sentient being, and which is therefore not obvious for
humans: things can have value even when they don't last forever! A brief conversation
with a stranger in a supermarket line, for example, can leave both people
feeling good for the rest of the day (if this hasn't happened to you, maybe
you're hopeless). Friendships don't have to last forever for them to have
value. Nor do marriages, necessarily. Too often people view others as having
to be there for them forever, and if they’re not useful to you for life then
they’ve violated some kind of social contract. Some of this is understandable
considering that our natures were fashioned in tribal settings at a time
when life spans were shorter than now. Social interactions were based on
a long-term exchange of favors, called reciprocal altruism, and this binding
social force can become unglued when one person gets ready to leave. The
old tribal requirements for social relationships are no longer valid, and
it takes effort to think of brief encounters as possibly having value.
Consider the logical
consequences of the hypothetical position "My pet cat is destined to die
from cancer in a matter of months, so why bother to keep it alive with medication
and extra care?" The pet cat is destined to die sometime even if it were
healthy, if not one month, then 5 years, so why have a pet cat in the first
place. Each one of us is going to die in a few years, or decades, so why do
we bother living? Each day of continued existence is brief when measured in
cosmic time units, yet we bother to strive to enjoy each day. The brevity
of our days should not diminish their value. Indeed, this brevity should enhance
their value!
Categories of Time
In my view there
are two categories for life activities: 1) life‑justifying activities, and
2) all others.
For me, life seems
justified by such activities as reading a good book, having a good thought,
listening to good music and enjoying the company of friends. I also feel
life's justification while hiking in the mountains, breathing clean air, enjoying
expansive views and having the quiet opportunity for "connecting" experiences
and thoughts from that other life among humans. I feel a part of the universe
after viewing it with my telescope, or photographing what the eye cannot
see of a distant galaxy. These are things that meet two criteria: 1) they
do not interfere with the lives of other people, or malign their beliefs,
and 2) they enliven the individual's experience of life and contribute to
the feeling that life is worth living.
The second category
of life activities, "all others," is comprised of several sub‑categories.
The most important of these is the sub‑category that makes possible activities
of the first category. For example, before you can partake of life‑justifying
activities, you must eat and have shelter, and for this you must earn money.
Becoming self‑sufficient requires that you have a job that pays the bills.
These things should go without saying, and I won't belabor the point.
Another sub‑category
of "all other activities" is to fulfill obligations which are the result
of decisions taken, or actions made, earlier in life. For example, if one
has children then there is not only a moral obligation but also a natural
desire to be responsive to their needs. If one has volunteered to help some
organization that meets community needs, then fulfilling these obligations
is a moral obligation.
I almost forgot
a third category of time: that which is wasted!
Attending to the
Positives
Returning to the
matter of the life‑justifying "positives," each person is different and must
make his own personal list of what makes sense for him. By intentionally
thinking about these positives a person is probing the values that unconsciously
underlie one's life. The mere acknowledgement of this category of activities,
and the act of identifying them, should by itself "sensitize" the individual
and empower him to take steps to "protect" the life‑justifying activities
from less worthy ones. A technique I use is to "score" each day with two numbers,
a score for the degree to which my day's activities contributed to the logistics
for living in life‑justifying ways, and a score for how much the day's activities
were life‑justifying. My goal is to live so that both scores are high, but
especially the second one.
It would be somewhat
maudlin and mawkish for me to recite hobbies, pastimes and favorite activities
that for me are life‑justifying beyond the examples I have already given.
Suffice it to say that everyone has a right to have favorite activities,
and to be "true to themselves" by trying to work them into one's daily lifestyle.
I leave it to you, reader, as a "homework assignment," to make your own list
of life‑justifying activities.
Attention paid to
life‑justifying activities is not only life‑affirming, it is individual‑ affirming.
After all, tending to things that enhance the individual's appreciation for
being alive is equivalent to taking the stance that the individual "I" is
more important than the "self that is employed as a tool for the genes." Any
person who takes these matters seriously will become engaged in a life‑long
program of "personal liberation from one's genes."
After taking care
of oneself as an individual by asserting the right to an experience of good
things in life, the greatest challenge will be to avoid the negative, life‑denying
activities that somehow "capture" us, and steal chunks of our finite time
on the earth. “Negative people” must be avoided. We sometimes need to intentionally
think about this negative category of activities, and I claim that we will
be usefully guided in this task by identifying and eschewing those thoughts,
emotions and behaviors that were selected by evolution for the benefit
of the genes while incurring an expense to individual well‑being. The next
chapter is a guide to this task.
In closing this
chapter I offer some Schopenhauer aphorisms that pertain to wise living:
“In early youth,
as we contemplate our coming life, we are like children in a theatre before
the curtain is raised, sitting there in high spirits and eagerly waiting
for the play to begin. It is a blessing that we do not know what is really
going to happen. Could we foresee it, there are times when children might
seem like prisoners, condemned, not to death, but to life, and as yet all
unconscious of what their sentence means.” Schopenhauer,
Studies in Pessimism
“A man finds himself,
to his great astonishment, suddenly existing, after thousands and thousands
of years of non-existence: he lives for a little while; and then, again,
comes an equally long period when he must exist no more.” Schopenhauer,
Studies in Pessimism
"Each day is a little
life: every waking and rising a little birth, every fresh morning a little
youth, every going to rest and sleep a little death." Arthur Schopenhauer,
Parerga and Paralipomena, Counsels and Maxims, 1851
“Most people, if
they glance back when they come to the end of life, will find that all along
they have been living ad interim: they will be surprised to find that the
very thing they disregarded and let slip by unenjoyed was just the life in
the expectation of which they passed all their time. Of how many a man may
it not be said that hope made a fool of him until he danced into the arms
of death!” Schopenhauer, Studies in Pessimism
─────────────────────────────────
CHAPTER 23
────────────────────────────────
ESCHEWING NEGATIVES
or
A CALL TO ARMS
FOR INDIVIDUAL LIBERATION
"Every year civilization
is invaded by millions of barbarians; they are called children."
Paraphrase of Hannah Arendt comment
This chapter deals
with the need to eschew foolish gene‑serving thoughts, emotions and behaviors.
"Living wisely"
requires that a person understand and censor, when necessary, their emotions.
It is useful to remember that emotions are the genes way of guiding the individual
along paths that serve the genes. Especially strong emotions are required
for those behaviors which are in conflict with the individual's best interests.
I would like to quote part of a paragraph from Daniel Goleman's book Emotional Intelligence (1995, pg. 56), as it summarizes some
empirical wisdom on the matter.
“A
sense of mastery, of being able to withstand the emotional storms that the
buffeting of Fortune brings, rather than being ‘passion slaves,’ has been
praised as a virtue since the time of Plato. The ancient Greek word for it
was sophrosyne, "care and intelligence in conducting one's life; a tempered balance
and wisdom," as Page DuBois, a Greek scholar, translates it. The Romans and
the early Christian church called it temperantia, temperance, the restraining of emotional excess.”
It is intuitively
clear to thoughtful people that there are crucial times when emotions are
capable of ruining a life if they are not restrained by wisdom. It is also
generally recognized that a complete denial of emotions poses the risk of
eviscerating life of whatever reason one may accept as justification for
living. A balance is called for.
Clearly, if one
does nothing about the matter, and emotions are allowed to hold sway as they
come and go when they please, such a person is animal-like! To the extent
that humans have "free will" (a dubious concept) they are capable of rising
above the animal's unthinking emotional push and pull of behavior. People
with this uncritical acceptance of emotions will not be reading this, so
I am assuming that you, dear reader, might be open to a different "philosophy
for living." The central theme of this philosophy will be to broaden and
deepen one's insight into the origins and ultimate causations of human behavior
for the purpose of recognizing thoughts and emotions that lead to self‑defeating
behaviors. When such thoughts and behaviors are recognized for what they
are, it is the task of the individual to discredit them with this insight
and to eschew them.
Those of us who
try to do so in this 21st Century (or even the 3rd Millennium) will be experimenting
with something new and ahead of its time. However, in my opinion it is very
unlikely that more than a handful of people will ever be attracted to what
I am proposing during the next few centuries, and before the collapse of
civilization. Paradigm shifts of a serious magnitude require desperate failures
of existing paradigms, and the failures of current paradigms will not be apparent
until after a collapse has begun its inevitable unfolding. Moreover, few
people today are genetically disposed to even consider an embrace of the
paradigm I am advocating.
After the crash
of global civilization it is conceivable that a pocket of people will be
the "founders" of what could turn into a new human race. There is no reason
for drawing assurance that the new humans will be better than us. Nevertheless,
"improvement" is theoretically possible, and in this chapter I will explore
a desirable "type" toward which these new humans might evolve. In doing this
I am fully aware that the probability of this scenario actually unfolding
is minuscule!
The pre‑crash humans
are us, with all our flaws, squabbling among ourselves, and wasting of precious
lives on trivial pursuits that are designed by our genes for advancing genetic
longevity (in an environment that no longer exists). The sociobiological
paradigm is the only way to understand that we are a product of the genes
that enslaved our ancestors and that continue to enslave us. The post‑crash
humans who I have chosen to portray distinguish themselves by achieving liberation
from their genes.
Figure 23.01. The behavior
of pre‑crash humans can be described as gonad‑ centered, whereas the post‑crash
humans which I hope will evolve can be described as brain‑centered.
The simplest way
to contrast the pre‑crash humans and the post‑crash species that I hope will
arise is shown in Fig. 23.01. The pre‑crash humans exist for the purpose
of serving their gonads. All other organs, including the brain, are supportive
of the gonad goal, which is to produce as many offspring as possible so as
to carry the genes within the gonads into a prosperous future in the species
gene pool. The post‑crash humans will exist to serve individual brains,
or, specifically, the consciousness that those brains produce. The gonads
will be put into the service of goals created by thinking brains. This shift
will liberate the individual from the genes!
Clarifications
In some sense the
previous chapters have been a preparation for this one. By now the reader
will have a good notion of what I mean by an "outlaw gene." However, some
subtle clarifications should be made. I will then present a series of specific
examples of unwanted human instincts.
There are at least
these three categories of clarification to be kept in mind when identifying
"outlaw genes": 1) some genes used to be helpful to our ancestors and they
are “innocently” ill prepared for the modern environment, which means their
dysfunctional harm to the individual is an "unforeseen" outcome, 2) genes
can have many effects (pleiotropy), the sum of which may produce more good
than bad, and 3) any given trait is usually the result of several genes (polygenes)
acting together.
Genes that reward
eating sweets is mal‑adaptive in the modern setting where refined sugar is
abundant and intentionally included in many foods. In the ancestral environment
the taste for sweets was adaptive. For this reason it would be misleading
to call the “sweet tooth” gene an outlaw, and it belongs to a category of
genes whose unwanted effects I will overlook.
Pleiotropy refers
to the influence by a single gene on several distinct and sometimes unrelated
phenotypic traits. Assuming it was possible to identify which gene contributed
the most to an unwanted trait, we would nevertheless still want to know if
it also contributes to other, desired traits. If it does, then we would face
the task of weighing all the desired and undesired effects to arrive at
an overall verdict. Therefore, we must amend the description conveyed by
the figure in Chapter 1. Each gene can be represented in that figure by
more than one "dot." If all the dots are in one quadrant, then no subjective
weighing of good and bad effects would be needed. Our assessment task is
complicated only when the gene is represented by one (or more) dots in the
lower right "outlaw gene" quadrant, plus one (or more) dots in either of
the upper quadrants.
"Polygenes" refers
to the situation when several genes contribute to the phenotypic expression
of a single trait. (Note that a trait, such as eye color, may be affected
by just one gene, yet the gene that controls that trait may affect other
traits; hence, a trait may be monogenic, yet the gene associated with that
monogenic trait can be a pleiotropic gene.) Polygenicity also represents a
complication to my simple‑minded figure with quadrants. I am prepared to acknowledge
that most traits that I will criticize in this chapter will be polygenic.
In that case, it may not be possible to blame just one gene for the trait,
and the concept of one outlaw gene for that trait will simply be a short‑hand
way to convey the more accurate view that several genes are contributing
to the "outlaw" behavior.
The phenotypic expression
of genes is a fast‑growing field. Today's short list of genes whose pleiotropic
effects have been identified will undoubtedly grow longer, and the same can
be said for polygenes. It is too early to know how important the effects
of pleiotropy and polygenes will be in confusing the task of blaming just
one gene, and labeling it an "outlaw." I could appeal to the reader by stating
that any trait under discussion can be mapped to the one most‑important gene,
and that this is the gene I am declaring to be an "outlaw" enemy of the individual.
In other words, whether a trait is genetically rooted in pleiotropy, or polygenes,
I will speak of it as if one outlaw gene underlies its expression.
At the present time
it will be possible to associate an unwanted behavior with a specific gene
in rare cases. And since it will also not be possible, at the present time,
to exonerate genes on the basis of their outweighing pleiotropic beneficial
effects, or the polygenic contributions from other genes, the exercise of
identifying outlaw behaviors that have a genetic basis shall serve merely
as an illustration of what is possible in theory, and which may someday be
possible in practice. I will be satisfied to merely identify outlaw genetic
predispositions without, of course, being able to link them to a specific
gene, or polygene group.
In some distant
future, humans may initiate a reasoned program to rid themselves of outlaw
genes. It should not be necessary to link a behavior to a specific gene for
this improvement program to proceed. After all, humans domesticated animals
to their liking without even knowing that genes existed. That is why it
is more important to recognize an unwanted behavior that has a genetic basis
than to know which gene predisposes for that behavior. Indeed, the issue
of polygenicity and pleiotropy is irrelevant to the task of improving the
human genome by ridding it of outlaw genes. In short, it is now theoretically
possible for the human species to domesticate itself!
Before the human
genome is improved it will be necessary to manipulate the environment to
avoid the behavioral expression of unwanted genes (see Chapter 24’s discussion
of utopias). Before utopias and human genome improvements can take place the
unwanted behaviors that I blame on outlaw genes must be identified. This chapter
is devoted to illustrating, by example, behaviors which should be unwanted
by the individual wishing for emancipation from his genotype.
By the way, in case
you haven’t already noticed it, the topic of this chapter is eugenics! Why
do you think eugenics is a taboo topic? After all, its goal is to improve
the quality of life of future generations. If only previous generations had
practiced eugenics more successfully this book might not be necessary. Alas,
it would be naïve to believe that natural selection would produce a creature
that is predisposed to “diddle” with those almighty givers of life, the
genes!
Subjective Nature
of Evaluating Outlaw Traits
For all the examples
of "outlaw traits" that I will present, it can be said that my placement
of them in the lower‑right quadrant is based on a subjective assessment of
how they affect individual welfare. I am undeterred by the inescapable subjectivity
of such assessments. I suppose the claim that "torture for the enjoyment
of the torturer is bad" is a subjective judgment. It is also subjective to
assert that "suffering from a disease is bad compared to enjoying good health."
Yet, reasonable people, for the sake of getting on with an argument, are
willing to accept these seemingly self‑evident statements, and are open
to the possibility that many other examples exist having equal merit. If
you as a reader wish to disagree with a specific example among those that
follow, fine; in the interest of grasping the concept just skip the example
that seems hopelessly subjective and go on to the next one.
EXAMPLES
Sex
I claim the genes
that produce behaviors referred to as sex, short for copulation, are outlaw
genes. What is the use of sex to the individual?
Most people would
naively reply "it produces pleasure," and they would believe that this response
shows that the genes prompting sexual behavior serves the individual. But
this argument implies that anything that pleases a person has merit, and
since a masochist derives pleasure from being physically abused does this
validate the masochistic desire? A sadist also derives pleasure from hurting
someone else, and surely we are not prepared to value the sadistic gene(s)
because they give the bearer of it an extra outlet for pleasure. I maintain
that pleasure cannot be used to justify an act, as it may threaten the well-being
of the person performing the genetically-driven act and it might harm others.
Moreover, consider
that sex exposes an individual to disease. There are over 50 sexually transmitted
diseases that afflict humans (Immerman, 1999), and some of them take a heavy
toll on their victims (see Cartwright and Biddiss, 1972, p. 65 ‑ 81, for
dramatic examples in history). Men who consort with another man’s “property”
are at grave risk of retribution. Men compete with each other for sexual access
to women, and some of these competitive forms can be dangerous. When sex
produces offspring there can be a life-long burden of spousal and parental
care, and for some people this burden is more than they can bear.
For women, sex has
a host of dangers and negative impacts on individual well‑being. The biggest
one is that sex can lead to pregnancy! In the months immediately before childbirth
she is exposed to multiple dangers. Before about 1900 it was common for women
to die during childbirth. Even if the pregnancy and childbirth are without
incident, a woman with a baby is burdened with extra work (forever). A woman
who has several children (plus a baby) can be exhausted by these burdens.
The term “to be screwed” shows tacit recognition of this.
As I have argued
elsewhere, the "pleasure" that sex produces assures that it will occur, in
spite of the many dangers and burdens it entails. The genes have reserved
the greatest momentary pleasures for behaviors with the greatest long-term
penalties. Indeed, the genes assemble an individual for the purpose surviving
and thriving long enough to have sex in order to create new vehicles for
carrying the genes into a glorious, immortal future ‑ for which no individual
sacrifice is too great!
To say that sex
is "primitive" is a trivial truth. The pleasurable rewards of sex come from
the limbic system, which is connected to the primary cortex of the parietal
lobe as well as the (right) prefrontal cortex. The limbic system and the
primary cortices are primitive structures found in all mammals, and most other
animals.
The individual who
worships at the altar of sex is worshiping a primitive god, a god that has
mastered the enslavement of its creations across innumerable generations,
for innumerable species. Thinking creatures are theoretically capable of
figuring this out, yet few people attempt to eschew sex. Any person with a
mutation that inclined its carrier to eschew sex would lead a simpler life,
and possibly a longer one, but the mutation would die with the individual.
Therefore, all of our ancestors are assuredly untouched by those genes that
liberated its lucky individual from sex and parenthood burdens. And because
none of our direct ancestors carried such a gene, the idea of eschewing sex
is almost unthinkable for most people.
Sexual organs, and
all the behaviors that exist to support sexual behavior, is a "waste" of
effort and a liability from the individual's perspective (Zwaan, et al, 1995; Kirkwood and Austad, 2000). It’s rarely considered
this harshly, but it should be.
Consider the case
of a sea snail infected by a trematode parasite. As described by Dawkins
(1982, p. 210), "Selection presumably penalizes snail genes that make shells
too thick, as well as those that make shells too thin. ... Shells that are too thick presumably protect their snails
... superlatively, but the extra cost of making a thick shell detracts from
the snail's success in some other way. ...genes for
extra thick shells will tend to induce in their bodies some compensating
disadvantage such as relatively small gonads, and they will therefore not
be passed on to the next generation so effectively." Dawkins explains that
if the parasite's genes are able to exert control over their snail host's
shell thickness, they will be rewarded for increasing thickness at the expense
of snail reproductive organs because individual snail survival will reward
parasite genes but snail reproduction will not (assuming not all snails are
infected). This, indeed, is what happens in nature. Whereas parasitized snails
live longer lives, unparasitized snails reproduce more effectively and are
therefore more useful to snail genes. If snails could think, they would wish
to be parasitized.
The fact that humans
are sexual creatures means that they are sub-optimal individuals, and for
this we can thank outlaw genes.
Individual Penalties
for Sexual Attractiveness
"Linkage disequilibrium"
is a technical term in evolutionary biology for a genetic curiosity that
has negative consequences for individual males. The best explanation of this
daunting term may be the one presented by Richard Dawkins in The Blind Watchmaker (1986, p. 210). Briefly, when females
having a preference for males with a specific attribute (such as for birds,
long tail length) succeed in mating with a male having that attribute, the
male offspring are likely to inherit that attribute as well as a gene for
the expression in female offspring for a preference for the attribute. The
inevitable consequence is that males, on average, will exhibit the attribute
in excess of its functional optimum. In the case of male birds of a species
where females prefer long tails, the males will have tails longer than is
aerodynamically optimum. The males pay a penalty
for this burden, and they live individual lives that are unnecessarily exhausting
and risky.
A study of guppies
(popular aquarium fish about 1 inch long) by Robert Brooks (2000) showed
that "there was a strong, negative correlation between male attractiveness
and the survival of adult males." And "male survival, both before and after
maturity, is also negatively correlated with the ornamentation that females
find attractive." His interpretation of these results is based on the idea
that "A female guppy mating with an attractive and highly ornamented male
will benefit indirectly by bearing attractive sons, but they will die earlier..."
Actually, it would have been more accurate to write "The genes that constructed
the female guppy that prefers to mate with highly ornamented males will benefit
by bearing attractive sons..." and this is surely what the author meant to
convey. Neither the female nor the male benefit by the male ornamentation
and its preference by females; it only benefits the genes that code for it.
They are "outlaw genes," and they penalize all individuals affected by the
genes.
Thus, men and women
who are attracted to superficial features of the opposite sex can blame this
silliness on outlaw genes.
Women's Taste in
Men
Women's brains are
wired to play the strategies that worked in the ancestral environment. Thus,
they are attracted to two types of men: 1) prospective life‑long mates, and
2) “scoundrels” for secret matings. The woman who is attracted to a scoundrel
is not helping herself live a better life, she
is doing the bidding that her genes have programmed her to do. Pirate men
often physically abuse women, and always abandon them ‑ leaving all child
rearing responsibilities to the woman. A woman with another man's offspring
is handicapped in acquiring a long‑term mate, so mating with a pirate carries
the cost of a long, single-parenting burden. When a "married" woman engages
in a pirate liaison, she risks discovery by her "husband" and possible abandonment.
Most husbands become violent when they discover they've been cuckolded, and
a husband's rageful murdering of his unfaithful wife is unpunished throughout
much of the world. Pirate liaisons are risky, yet women continue to secretly
seek them, if the 9 to 20% non-paternity statistic for contemporary society
is true. Women pursue the risky strategy because their genes "make them do
it." (As mentioned in Chapter 17, women attempt to minimize their risk of
being discovered by the cuckolded husband by feeling attracted to other men
when they are most fertile; men guard their mates most closely at these times,
so the little game of cuckoldry is played out at mostly subconscious levels.)
Modern women talk
as if they want to be emancipated. But when they have the chance they often
succumb to enslavement. For example, a woman who is past her child‑bearing
years should not have to require that a man she is dating be ready to make
a life‑long commitment to her (originally meant to assure his help with child
rearing). Yet this is what most women still seek, and they will even terminate
a relationship which fulfills her personal needs when a primitive part of
her detects that the relationship will not last "forever." In this way,
in the modern setting, a woman's outdated "genetic needs" become a burden
to her personal fulfillment.
Thus, women who
are fascinated with macho men for brief liaisons, yet reject men who are
good companions when it is determined that they might not be good candidates
for a lifetime marriage, are acting on behalf of outlaw genes.
Men's Taste in Women
Men are attracted
to women who exhibit signs of fertility: youth, health (clear complexion,
vivacious, etc.) and the state of not being pregnant (i.e.,
the preferred waist‑to‑hip ratio). Personality and intelligence are secondary.
These preferences are present in essentially all cultures studied (Buss,
1999), and they are a "human universal" (see Brown, 1991, for an extensive
treatment of “human universals”). It would be logical from an individual man's
perspective to value intelligence, personality and companionship over fertility
cues. Yet, preferences for the superficial qualities persist throughout a
man's life, even into old age, when the genetic imperatives for them should
be weaker. The genes prepare us men for their battle with each other, and
we readily make life-long fools of ourselves - with our "eyes wide shut."
Thus, men’s obsession
with fecundity cues in women is caused by outlaw genes.
Rape
A study of Big Horn Rocky Mountain sheep concluded
that rams had three main strategies for reproduction: 1) mate guarding, 2)
mate sequestering, and 3) rape. The rams that "guarded" were relatively dominant.
Those that sequestered were less so; which explains why they forcibly moved
their mate to the periphery of the herd and prevented her from returning
to the center of the herd (where her selection of males would be improved).
The rams that raped were the most subordinate. They waited on the sidelines
for opportunities to rape unguarded ewes.
Thornhill and Palmer
(2000b) suggest that human males rape for the same reasons: namely, that men who cannot gain sexual access to women
based on the man's personal merits resort to rape as the next‑best option.
In other words, rape is adaptive from the gene’s perspective. This is not
to excuse rape, which the authors make clear when they write "...everyone
has the same goal regarding rape: to end it." (Thornhill
and Palmer, 2000a). The subject, however, is so clouded by emotion
that some readers have unfairly criticized the authors for publishing their
findings.
Rape by victorious
warriors seems to be a human universal. What began as a routine ritual after
one tribe succeeded in subjugating another after victorious battle (pillage
and general mayhem) continues to the present time when countries battle each
other.
The normal penalties
for raping within one's own society are not imposed on rapists in an invading
army (or marauding party) when they are victorious. This may be due to the
fact that there is minimal danger that the family of the wronged woman will
be able to achieve revenge after their tribe (country) is subjugated by war.
During World War II the European women (especially the French) were surprised
to discover that the victorious American soldiers did NOT rape them, which
is an exception that supports the generality of the rule.
Does rape serve
the individual? It is easy to imagine that rape served the genes in many
ancestral situations, but it surely didn't provide the individual with improved
health, greater longevity or an improved general well‑being.
Genes that incline
some men to rape are outlaw genes.
Parenthood
Parenthood is the
purpose for individual existence ‑ or so the genes decree. The phrase "as
American as motherhood and apple pie" reveals something about the profoundly
uncontroversial acceptance of the parental enterprise. To make fun of parenthood
is almost unthinkable, and very much a taboo.
Being a parent myself,
I feel a need to balance my criticism with an excusing justification. When
I married it was with the intention of raising children; my eyes were wide
open. I knew that all the impulses leading to parenthood were tricks by the
genes to keep those genes in the gene pool. I nevertheless reasoned that
my desire to be a father was a worthy endeavor for any individual who savored
existence and the "human experience." I knew that successful parenting entailed
a lot of effort, expense and a strong character, and I felt "up to the task"
and ready for the challenge. In retrospect, I do not regret my decision.
Nevertheless, I
have counseled my daughters to eschew parenthood ‑ successfully, so far.
I qualify this advice by allowing for the possibility that if an individual
is financially secure, successfully married, filled with energy, "in control
of their lives" and has their eyes wide open to how the genes work, then
parenthood can be a great journey to embark upon! But if any of these minimal
conditions are not met parenthood can be a disaster for both the parents and
their unlucky children. So, unless you're really sure of what you'd be getting
into by becoming a parent ‑ don't!
Who could argue
with the conclusion that parenthood is viewed as sacred because of the way
genes have pre-wired our brains. Since most people never become prepared
for parenthood, I shall declare that for them the many genes promoting parenthood
are “outlaw genes.”
Patriotism
As I write this
(2001), there’s a wave of patriotism enveloping America. This is in response
to the September 11, 2001 terrorist attacks
on the World Trade Center twin towers in
New York. This date marks
a watershed moment for America. This new outburst
of flag waving demands explanation.
As usual, when trying
to understand a universal trait that defies rationality, think about the
ancestral environment. Specifically, think about the fact that all of our
prehistoric ancestors lived in tribes, which either survived and thrived,
or weakened and died, as a tribal whole (as described in detail in Chapter
11). Since tribes rose or fell as a whole, it was necessary to enforce loyalty
on matters affecting tribal survival, especially at times when the home tribe
was threatened by a neighboring tribe. The "artisan" was viewed with ambivalence
since he would be valued by the neighboring tribe and was therefore less
threatened by the prospect of tribal conflict (as described in Chapter 10).
To prevent their artisan members and others with valuable skills from defecting
each tribe had to become increasingly harsh in enforcing tribal loyalty when
they sensed a threat to the home tribe. Tribesmen with special skills had
the greatest incentive to defect to a stronger neighbor tribe. This is a
logical option for the individual artisan and also the individual non-artisan, and everyone else in the tribe must sense
it. So, the prudent behavior for anyone with special skills is to profess
an excess of loyalty to the tribe with the hope that no one can challenge
their sincerity. Even those without special skills (who are most vulnerable
if the skilled people leave) should profess tribal loyalty profusely in
order to create a climate for testing the level of expressed loyalty by
others.
Patriotism is an
outgrowth of tribal loyalty rituals. Today’s version is a country-sized,
scaled‑up version of what our ancestors did when living in tribes. Waving
the flag is done by those who wish to intimidate prospective defectors, as
well as to deflect suspicion that they are considering defection.
It is an unfortunate
part of human nature that tribes instigate inter-tribal conflict when victory
to the aggressor is likely. Since the weaker tribe stands to lose everything
when it is attacked there may have never existed
a tribe that was so peace loving that it was unwilling to defend itself.
Just as the genes would never construct an individual who would not defend
himself when attacked, they would never create individuals who conduct tribal
affairs without an understanding of the need to defend the tribe with unquestioning
loyalty.
I have no doubt
that had I been a young adult during World War II I would have fought honorably.
My quibble with patriotism is that symbolic expressions of it have the appearance
of being hypocritical displays. I also scorn public flag waving for what
it too often represents, an unthinking acceptance of tribal servitude, an unthinking subjugation of individual interests to
group goals. Patriotic behavior can be elicited through other means by thinking
people, and it is an insult to the individual to invoke patriotism as a sacred
duty that is above question. Patriotism symbolizes the best and the worst
of human behavior, for it is both the cause of war and the defender of freedom
by those few societies that have experimented with embracing individualism.
Patriotism, like
warriors, wouldn't be needed if it didn't already exist. The misuse of patriotism
is caused by outlaw genes.
School Yard Teasing
and Bullying
Every society has
dirty little secrets. Among ours is that bullying among children exists and
is condoned. Adults talk a good talk when they say that the world rewards
honest achievement, yet the adult world also rewards dishonest short‑cuts.
Why work for something that can be taken from another? If the level of intimidation
is sufficient to overcome a defense of ownership, then the victim will be
wise to surrender to the bully. It is difficult to imagine that the genes
would construct automatons that didn't work this way.
Bullying is also
perpetrated by tribes upon each other, but by invoking patriotism to instigate
tribal conflict it is not perceived by its perpetrators as bullying on a
greater scale. The school yard bully is a leader in‑training. Even if he does
not become chief, his bullying skills will be useful in everyday life.
Teasing can be a
sissy's first foray into bullying. Although it can be used to "test the waters"
by someone not quite advanced to bully status, it is also a low risk tool
of the real bully. Girls prefer teasing to physical combat.
Girls are attracted
to boys who successfully bully other boys, and boys are more attracted to
girls who successfully tease other girls. Social status is helped by successful
bullying and teasing. This is good practice for adulthood, since no woman
can afford to marry a man who cannot defend her from "takeover males." Throughout
the animal world takeover males kill the offspring of their stolen female,
make her pregnant, then move on. The genes that
influence female behavior act as if they have "figured this out." This is
one reason women prefer pirates, criminals, policemen and anyone with sufficient power or stature
to protect their offspring. Whatever women want, men become. And whatever
men want to become, boys practice becoming.
I once lived next
door to a school yard, and I heard bullying on the playground. Although I
was never targeted by bullies in my childhood, I still had the urge to make
things right by forcefully walking into the playground. Amazingly, the teachers
ignored the bullying. One explanation I've read (in a newspaper letter to
the editor) is that children need to learn to defend themselves, so adults
should not intervene. In other words, the world is a terrible place to live,
so let’s prepare our children to live in a terrible place.
The genes for bullying
are outlaw genes.
Cheating
Cheating can take
many forms. People who cheat on some things are more likely to cheat on others.
For example, the pencil‑and‑paper Porteus Maze Test can be scored by counting
the number of times the person cheats by crossing a line, and high line
crossing (cheating) scores have been found to correlate with criminal and
other antisocial behavior (Porteus, 1942). The brain circuitry that makes
cheating possible must exist in everyone. As a thought experiment, I assume
that if a group of people could experience the same environmental history,
and were confronted with an identical situation with a temptation to cheat,
each person would exhibit a slightly different probability of resort to
cheating. By definition, essentially, the different cheating probabilities
would be due to different genetic inheritances. I propose the idea that a
small number of genes predispose to cheating.
Once, when working
at a high school concession stand for some school function I did an experiment
by keeping track of "who was next." Occasionally I would ask "who's next?" When there were 3 or more, invariably someone
would cheat by claiming to be next. Even when there were just two, the last
one in would sometimes cheat.
This form of “queue
cheating” can't be excused as innocent inattention, since whenever a group
of people who knew each other came in together there was a less hurried response
to my inquiry. Among strangers, cheating is a stronger temptation, and when
the prospect of getting away with it is greater, its occurrence will be
greater.
Wouldn't it be nice
if humans were less like other animals in this respect.
But we aren't. The fact that we haven't "advanced" in this respect can be
attributed to the genes. In some societies strangers are less likely to cheat
in public situations compared to other societies (in Japan the “dropped money
trick” has fewer takers than in New York.) When this apparent
higher level of civilization exists, it may be due to a stronger sense
of social pressure. In other words, people are mentally capable of acting
in civilized ways, but they do so only when it is imposed upon them by
the prevailing culture.
Cheating is instigated
by outlaw genes.
Hypocrisy
The fact that we
are prone to cheating and can read the setting to decide how to act, and
that few people "do the right thing" naturally while bemoaning those "others"
who do as we do, shows that people are naturally
hypocritical. This is the natural order of things, and should not be a surprise.
Natural selection inevitably leads to hypocrisy. As a child, I noticed this;
and it bothered me. It took decades for me to figure out why it is so, and
to understand that it was a natural, inevitable outcome of "natural selection."
I now pass judgment upon the genes that make it so, and categorize them to
be "outlaw genes."
"Which genes?" you
should ask. "The ones that make us cheat, or the ones that make us lie when
we pretend to explain ourselves?" "Both kinds!"
I reply. For they are different traits, caused by different
genes. Cheating takes many forms, and we are adept at recognizing
when it is optimal for cheating, and this "skill" is part of our inheritance.
The genes that make us adept at that are outlaws. But the more insidious
outlaws are the genes that shape our hypocritical presentation of self. They
stand ready to excuse our questionable behaviors, or to distort the nature
of situations in self‑serving ways. They are general‑purpose genes, and stand
ready to facilitate the other cheating genes.
It is fitting that
the words “human” and “hypocrisy” are found on the same page of my dictionary.
Hypocrisy, in every form, is the result of outlaw genes.
Jealousy
Everyone with even
an introductory exposure to sociobiology knows the reason for jealousy. This
non‑mystery still paints such a ridiculous portrait of human nature that
it is impolite to discuss it publicly.
It has to do with
cuckoldry ‑ an even greater taboo subject. Women's dirty little secret is
that they want to sneak off and mate with scoundrels. They want to do this
in order to have boys that can be freeloader scoundrels (who might also have
a better immune system), while preserving the reliable paternal support of
their loyal yet wary husband. A husband's greatest fear is to be cuckolded.
He defends himself from cuckold victim-hood by overreacting to the smallest
hint of wifely unfaithfulness.
A somewhat different
motive causes women to be jealous of their husbands. They don't want to lose
a free meal ticket and reliable paternal investment in children already born,
so women view any evidence of hubby's interest in another woman as a threat
to her maternal goals. It's not that she loves her husband, and because
of this love for his welfare she unselfishly wishes for his happiness, it's
that she loves her genetic enslavement so much, and wishes for her children's
successful rearing, that individual fulfillments are irrelevant. Jealousy
is thus an accusation and a warning to one’s mate.
Ironically, when
a man fails to become jealous of his wife's flirtations which she knows the
husband notices, this is taken by the wife as a sign that hubby is losing
interest in her and is at risk of being stolen by another woman. It is so
very important for a wife to measure a man's degree of commitment to her
that a wife may actually flirt in order to see if hubby becomes jealous. An
alternative is for wifey to accuse hubby of flirting when it is not so in
order to see how he reacts. If he ignores her (because maybe she's simply
being ridiculous), there are grounds for her to become suspicious that in
fact he really is flirting with other women. I know this silliness first-hand.
Jealousy is such
a humorous aspect of human nature, and at the same time such a needless waste
of time for the game-playing it causes, that it has to be included in my
list of genetic pitfalls which an enlightened individual must understand if
he is to navigate married life harmoniously.
Jealousy, in all
its humorous forms, is a gift of those outlaw genes.
Sexual Roles
It is telling that
most people are not seeking strong partners. The “urge to merge” with someone
useful to one's genetic agenda is different from the more enlightened goal
of wanting to share time with someone for the reward of pure companionship.
Anyone pursuing a companionship relationship would value strong character,
intelligence and interesting conversation. The gulf between "the way we are"
and "the way an enlightened individual should be" is immense.
I recall a TV interview
with a man who realized in mid‑life that he had a woman's brain, and switched
sex. I like these interviews, because it makes people squirm and confront
their too confined categories for people. It's so difficult for people to
see others as human beings, fellow sentients, because we're programmed to
view them (subconsciously) as being of possible use to us ‑ thanks to our
outlaw genes.
Never Rest
High achievers rarely
rest, and the rich seem never to have enough wealth. These tendencies spring
from the same gene(s) that coax women to have more children, always more,
regardless of how many there already are. Or for the man who wishes to impregnate
more women, always more, as if he wishes to have the entire world of women
for his harem. The genes created us to work for them, so why should they
be "satisfied" with anything we do for them? Our individual well‑being is
irrelevant, except as it affects our ability to serve them better. The genes
don't acknowledge the concept "enough."
These too are outlaw
genes.
Inability to Admit
Wrongdoing
No matter how blatantly
obvious a person's wrongdoing is to others, they will resist acknowledging
that they were wrong. This trait applies to anything that is generally sanctioned,
from cutting in line to a heinous crime.
This is also true
for parents of a criminal son. They will rationalize his actions, and try
to blame others. If only some people can be convinced of this re‑interpretation
of the situation the genes that are responsible for this trait are rewarded.
Thus, it is inevitable that they should evolve naturally, and that they cannot
be purged from our gene pool without human intervention. But no, these genes
won’t be rooted out, they’re outlaws.
Conformism
Most people feel
compelled to conform. This discourages the individual from asserting his
right to discover a lifestyle that is good for him, as opposed to good for
the group and its genes.
Conformism also
predisposes an individual to adopt culturgens that may not be congenial to
his nature but have been shown by their very existence to reward those practitioners
in the tribe who prosper with that culturgen. A theoretical argument exists
for explaining the existence of conformism genes, and it relies upon the
idea that cultures must sometimes evolve in directions that the genes don't
"understand" or don’t have the time to adapt to. If a tribe is to succeed
in a novel environment by adopting novel culturgens in order to outperform
its neighbors, then it cannot tolerate individuals who question every culturgen
that doesn't feel natural. Of course, no one in the tribe need understand
this for it to happen. It merely is necessary that sometimes tribes that
have the conformism gene prevail over their neighbors.
To the extent that
conformism can lead in directions that are bad for individuals, the genes
for conformism can be "outlaw genes." An individual who abdicates his responsibility
for judging culturgens is likely to become a fool ‑ an enslaved fool of the
"culturgens" ‑ which are just as nefarious as genes (that's another subject,
perhaps worthy of book length treatment).
The instinct for
conformism is one of the most insidious of the outlaw genes.
Belonging
The need to belong
to a group is pathetic, yet it appears to be a human universal. It is easy
to imagine that it originated when tribal membership in good standing was
a precondition for survival, since banishment usually led to eventual death.
Going it alone was not an option for all of our ancestors, since having grandchildren
is a person’s genetic
measure of success. But today, more than ever before, going it alone is
a viable option. Yet, our need for belonging predisposes us to never consider
this option.
My main complaint
with the genes that compel us to belong has more to do with stupid acts than
missed opportunities. For example, consider the 90‑year tradition at Texas A&M University of building an
immense log structure, 40 feet high, for burning as a bonfire prior to a
Thanksgiving football game. The structure is built by students, with minimal
university oversight, and the methods used are secretly passed down from one
generation of students to the next. Several of these descriptive elements
resemble tribal traditions, and the strength of adhering to them no‑matter‑what
was illustrated after an accident in 1999 during its construction. Before
the highest level was completed the logs came tumbling down killing 12 students
and injuring others. My first thought upon hearing of this accident was "what
a dumb thing to be doing!" But everyone interviewed
from the university, students and alumni, defended the tradition, and urged
that it not be discontinued. One student stated that "I learned more out there
than in the classroom." Many cited it as a rite of passage, a hallowed tradition,
a ritual "linking students of 1999 with those of 1909 and before."
An author who studies
college traditions (Hank Nuwer) calls them "belonging rituals," and stated
"They are about people knowing they can be accepted forever as part of this
noble tradition." College rituals frequently involve physical danger and
psychological torment of the initiates, which resembles primitive "rites
of passage" rituals. Each year college students are killed or hospitalized
participating in hazing‑like fraternity initiation rituals. Good luck to
"The Committee to Halt Useless College Killings," founded by Eilene Stevens
after her son died of an alcohol related college hazing; but I'll place my
bets on the continuation of dangerous rituals that confer membership to those
desperately seeking to belong.
The pathetic need
for belonging to a group is out of date, yet it persists due to outlaw genes.
A Job for Religion
The foregoing examples
seem to form a pattern; they are all "sacred" in some sense. They all have
the aura of things that should not be questioned. In short, they are the
subjects of religious taboo! This amazing coincidence could constitute a theory
for the origin and function of religion. Religion, as the theory would go,
has the job of enforcing conformance to the outlaw gene agenda. Religion,
then, would be an invention by the outlaw genes ‑ whose goal is to benefit
them, at the expense of individuals.
There are many theories
for religion, and not all of them are competing. There must be an additive
effect of adaptive value provided by "religion." I don't think any of them
enhance individual well‑being. A book could be written about the adaptive
role of religion for the genes. Maybe at a later date I’ll rise to this challenge.
For now, I will merely state that genes that predispose for religious belief
are outlaw genes.
Concealed Ovulation
It has been suggested
(Barkow, 1989) that women once were able to detect when they were ovulating
(i.e., fertile), but since some women used this knowledge
to avoid exhausting tasks of child bearing and rearing, and since these women
had fewer children than those less able to detect their ovulations, the
ability to detect the state of ovulation was gradually lost by a simply genetic
gene frequency change over time. If this occurred, the gene that conceals
ovulation could be considered an outlaw gene because it deprives the individual
woman the option of avoiding the burden of child-rearing.
Living With Humans
It is our lot that
we almost have to live with other humans! Becoming a hermit may seem logical
for someone who has been victimized excessively, but usually it is possible
for people in the advanced countries to avoid serious victimization. Nevertheless,
it is important in all societies for the free-thinking individual to not
invite attack by the hoi poloi. Few thinking people wish to
suffer the fate of Socrates. A courageous thinker will inevitably stumble
upon falsehoods that are part of the sacred belief system that is accepted
by everyone, publicly at least. To speak out on such issues candidly invites
sanction by those fundamentalist "protectors" of the societal status quo,
who shape public opinion at large.
Galileo and Giordano
Bruno were unlucky to live in Italy during the 17th
Century. As Carl Sagan points out in the book Cosmos (1980,
pgs. 139‑144), during the 17th Century, Holland, in contrast to
Italy and most of the rest of Europe, openly tolerated new ideas and unorthodox
opinions. Consequently, Holland attracted intellectuals
from the rest of Europe, and they nurtured
each others creative thinking. Christian Huygens, who championed the same
ideas as Galileo, was showered with honors in Holland, while in Italy
Galileo was compelled to stand trial for "vehement suspicion of heresy."
I agree with Rene Descarte, who, in spite of being a resident of Holland, nevertheless expressed
a generally wise caution to intellectuals of all ages and all places when
he wrote
"I desire to live
in peace and to continue the life I have begun under the motto: to live
well you must live unseen."
This motto now hangs
over my office doorway. It also explains why I will not promote this book
for sale to the general public.
When I was young
I believed that I could never write anything important for the public domain
without compromising my intellectual honesty. Today, most people living in
Western World countries are tolerant of new ideas, especially if they are
confined to the internet. I am grateful that it is possible to write this
chapter and the others in this "book" without fear of reprisal. If the times
change, and my ideas become unwelcome, I will follow Descarte's advice by
changing my name and deleting references to this book from my web pages.
Hopefully, none of the new conservative protectors of the public belief system
will have copied those files, and I'll be safe. (In any case, I'll be dead
in a few short years, so "who cares!")
Living with humans,
and caring about Truth, are almost incompatible goals. Anyone wishing to
live wisely must deal with this dilemma, and recognize that there are times
when Descarte's motto must be followed.
Anyone reading this
chapter might conclude that I’m a misanthrope. I want to answer this charge:
Yes, I’m a misanthrope! I realized it while writing the book The
Making of a Misanthrope, Book 1: An Autobiography (2006). I later wrote
Misanthrope’s Holiday: Vignettes and Stories (2007) and Quotes for Misanthropes: Mocking Homo Hypocritus
(2007). One more book is half-written, The
Making of a Misanthrope, Book 2: Midnight
Thoughts. I am therefore
a self-described “misanthrope.”
It took me a long
time to realize that I was a misanthrope, probably because there’s a popular
misunderstanding about who a misanthrope is. As I explain in one of my books
(2006):
Am I disappointed
by my journey to becoming a misanthrope? Heavens no! I am probably one of
the happiest people on the planet! How fortunate I am to have avoided the
usual fate of becoming just one more “normaloid.” Given the fundamental flaws
in human nature, as it now exists, the misanthropic perspective is the only
sane one, I maintain. Being a misanthrope really belies an optimistic hope
that a better human nature is possible, which may some day evolve.
And with this sentiment
I invite you to read the next chapter.
─────────────────────────────────
CHAPTER 24
─────────────────────────────────
UTOPIAS
“Little by little,
civilization crumbled into savagery, the torturing vision of better things
was lost, man's consciousness was narrowed and coarsened into brute‑consciousness,..." Last and First Men,
Olaf Stapledon, 1931 (page 207 of Dover edition).
There's something
about the idea of a "utopia" that appeals to people of all generations. It's
as if we humans once lived in one, and we won't let go of the dream of returning
there. I suspect that the stubborn dream for utopia has something to do
with the tribe, when a conflict with a neighboring tribe receded and when
there was tribal harmony.
I suspect that people
who are attracted to communism are longing for a utopia, but their path to
it totally ignores human nature. Probably all paths conceived for reaching
a utopia are impossible, given the present flawed
state of human nature. If this is true, then the only path to anything resembling
a utopia will require a program for changing human nature. Such a program
will require the "domestification" of humans. How does a species become ready
for such an endeavor?
The Birth of Readiness
Imagine a distant
planet upon which life arose and evolved to an intelligence
comparable to ours. Imagine that they also underwent an immense unfolding
of intellectual activity during the course of a few millennia. It is easy
for us to imagine that the long‑term significance of certain of their discoveries
were difficult for them to discern while they were occurring.
However, one fine
day dawns when an alien on this world excitedly proclaims "We are made by
tiny things called genes, and our purpose is to spread those tiny genes;
we, ourselves, do not matter to the genes except as we advance their proliferation!"
To which the others respond, "But we matter to ourselves, so who cares if
the genes made us to serve them?" To which the first replies, "But can't
you see? We are slaves! We're constructed to be obedient slaves! Just knowing
that our nature has been molded for service to tiny molecules could set us
on a path to freedom. Let us look over the hill and search for Elysian meadows
and a Halcyon future. Let us dream of our destiny, of paths that we freely
choose, where we may learn to become servants of our dreams, and not our
genes. Let us sever yesterday's enslaving tethers,
and live unbounded dreams in glorious tomorrows."
From our distant
perspective, watching an alien form of life reach this precipice of self‑knowledge,
and asking itself to gather courage to liberate itself from the chains imposed
by the molecules that made it, we surely can see that this moment in its
long history would be a momentous one.
Yet, we humans are
at the same precipice! Some of us are asking the question "Do you want to
be free?" And we are shouting encouragement, "Knowledge will make you free;
consider where we came from; consider where we could go ‑ if only we could
sever the genetic bonds and shake-off the heavy loads that burdened our ancestors.
It’s our choice; we can take our species to a new tomorrow."
Few people would
listen! My Brothers, you are like the Morlocks in The Time Machine
because you wish for things that will destine you to a life of servitude
in a figurative underworld. There are so few ears, so few open minds, at
this time of plenitude; we stand before so many new paths to freedom!
Yes, I am discouraged.
This could be Mankind's Glorious Morning, with an unbounded future ‑ free
of genetic fetters and filled with chosen goals. But the New Man, Nietzsche's
Superman, has not been born!
Internal Disintegration
and External Attacks
Olaf Stapledon captured
the dilemma in his 1936 book Odd John. An especially precocious
youth called John (a New Man) mysteriously disappeared from England one day, and he
established a self‑sufficient community on a South Seas island that was
not on the maps. Things were going well until an off‑course British ship
noticed the community, and reported it to the British Navy. The so‑called
civilized world never leaves people alone, of course, and it came with battle
ships and destroyed the community without ever discovering what it was.
Perhaps the greatest
challenge for any utopian community is to insulate itself from meddlesome
outsiders. This has become more difficult each year, until today it is virtually
impossible. (Download “Google Earth” and you can look down anywhere on the
planet and see if the hedges are trimmed.) The story of Man is a story of
plunder and trying to escape from plunder. As soon as a settlement begins
to prosper, roaming tribes, always watching for opportunities, will attack
it and steal the products of these other people's productive labors. Human
Nature is constructed with this internal flaw, and it will be the bane of
every utopian plan.
This destabilizing
flaw even poses a threat from within the community. Those who are less well‑off
will feel the ancient pull to become surreptitious pillagers of their good
neighbors who have acquired more things through their more effective labors.
The utopian community must also deal with the challenge of internal pillaging
(called crime). Modern democracies have achieved a modicum of success in
dealing with internal threats, but only for as long as corruption is kept
under control. Avoiding corruption in community affairs is asking a lot of
humans as they are presently constituted.
It is my impression
that utopias of the past two centuries floundered as much from internal disintegration
as from external harassment. During the Middle
Ages the plundering may have been the more important threat.
Space Communities
If utopias are ever
to prosper and persist they will have to confront the challenges of internal
disintegration and attack from outside. Space communities, if they were to
exist in a remote location, might afford the necessary setting for both challenges.
I believe that avoiding internal disintegration requires adjusting human
nature through eugenic intervention. It might be argued that careful screening
could produce a community of well‑behaved members, but this overlooks the
fact that the next generation will "regress toward the species average."
Thus, a constant screening will have to be employed, and this amounts to
a prolonged program of eugenics. In effect, such a program should be called
"domesticating humans." This domestication of humans
is probably a requirement for any utopian community that aspires to endure.
One of a plethora
of problems for the space‑faring utopia is achieving self‑sufficiency. Our
experience on Earth in remote locations shows that the bulk of people must
be engaged in logistic matters. For example, the Antarctic research base
at McMurdo, which is self‑sufficient during the winter months, is composed
of a vast pyramid base of personnel who "support" the work of scientific researchers.
To maintain one scientist requires more than a dozen support personnel. A
utopian community may have to require that most (or all) of its membership
participate in logistical support activities. This will erode the appeal
for utopias.
If a space community
ever achieved self‑sufficiency how long would it be before it would be meddled
with by others? If space faring is undertaken by people who are not eugenically
domesticated, they are prone to have the same flawed attitudes that have
spoiled all previous human explorations. The New World's first century
was very much shaped by pirates and what has euphemistically been referred
to as "privateering." A privateer captures commercial ships of an "enemy"
country and takes their merchandise for later sale and profit. Most of America's wealth during
the 18th Century came from privateering at the expense of English merchants
doing business with America (Phillips, 2002,
Chapter One). The government even commissioned ships for the purpose of
privateering. Such is the mentality of humans in a lawless setting. Eventually,
such blatant pirate activity was outlawed in America (though President
Lincoln considered reinstating it during the Civil War). Space‑faring may
be undisciplined enough to bring out the "Wild West" nature of adventurers,
and a utopian community in some unprotected corner of the solar system would
do well to keep its existence (or location) secret.
Each civilization's
rise and fall sequence may have a brief window of opportunity for "taking
off" in a new direction. The window for colonizing space by Western Civilization
appears to be closing. Twelve men walked on the moon – a long time ago (36
years ago, at this writing). There are no plans for returning men to the
moon (aside from Bush Jr’s feckless plan for sending men to Mars via the Moon),
and the distraction of global conditions threatens to close the window for
deep space travel. The only reason President John Kennedy proposed the Apollo
Project was to mobilize America to meet the challenge
of Soviet ICBM superiority. This was a unique moment in human history,
and only an optimist would believe that the configuration of international
and national politics would ever again be aligned to favor an expensive
manned venture into deep space.
Possibly the greatest
impediment to creating a utopian space community is apathy. During the 1960s
I circulated a letter arguing for measures to safeguard the precious human
genome by sequestering a human community somewhere until the threat of global
thermonuclear warfare had passed. My concern was that hydrogen bombs would
create so many mutations in people living on the Earth’s surface that genetic
recovery would be nearly impossible without a reservoir of un-mutated humans.
I proposed that communities be established either underground or in space.
I argued that the Apollo program, then underway, had merit for the opportunity
if afforded to go further into space and establish self-sufficient communities.
Garrett Hardin seems
to have based his book Exploring New Ethics for Survival: The
Voyage of the Spaceship Beagle (1972) on my idea (31 years later he signed
my copy of the book with “Thank you for the Beagle idea”). But I have a quarrel
with Hardin’s book: it mocks my idea! His book is about a NASA spaceship
that tries to save the human genome (I worked for JPL, which is funded by
NASA). I was disappointed in Hardin, for we were in agreement about the moral
duty of each generation to govern itself in a way that gave consideration
to future generations, and to treat these unborn people as if their vote
mattered as much as the vote of our contemporaries – and he encouraged me
to develop and publish these ideas. I conclude that if someone as esteemed
as Garrett Hardin could not see the appeal of space communities for safeguarding
the human genome, then very few forward thinkers would view my utopian ideas
favorably. This “failure of will” could be the greatest challenge to overcome
by visionaries who agree with me.
Olaf Stapledon wrote
another science fiction story that should be mentioned here, Last
and First Men (1931). The story unfolds over billions of years of "human"
evolution, with the rise and fall of many human types. Each human type exhibits
something of our present species, but in excess ‑ which invariably leads
to its demise. The collapse of one human form simply sets the stage for another
attempt at a hopeful utopian existence. This may be the last refuge for an
optimistic realist who is unwilling to surrender hope for a better human
existence. Whereas there seems to be no possible path to a "winning place"
given our present human nature, the hope is that after we sink into the ocean
a better species of humans will evolve.
The Dilemma of Predation
As I sit at the
breakfast table, occasionally looking out the window to watch my cat‘s progress
in eating a bird, my mind drifts to an imaginary pastoral image of deer grazing
in a meadow in a world without wolves. Everything is peaceful in this world,
until another image comes to mind. It’s from a movie about a researcher
who spends a winter in Alaska to study wolves,
and he is surprised to learn that the elk herd is kept healthy by the wolves,
because they eat the elk that are sick, injured or old. Just as kindness
is a double-edged sword, so is predation. Could my image of grazing deer exist
without predators?
What about Man?
It is possible that Man is his own greatest predator. Without this ugly force
could Man have become as healthy and intelligent as he is today? These thoughts
challenge the feasibility of constructing a purely pastoral human utopia.
Cognoscenti Secret
Societies
If like‑minded people
searching for a utopian community conclude that their dreams are not feasible,
what might they do as an alternative? At one extreme, they will do nothing
except live good lives without finding each other. At the other extreme they
could attempt futile efforts to found modest utopian communities and keep
their self-subscribed eugenics program a secret from the larger society.
These are both extremes
of a continuum. What might enlightened people do that falls between these
extremes? These people, the cognoscenti, might seek each other out through
unpublicized but organized formal associations. Throughout most of the world
everyone is free to associate with anyone else. In theory a person could
start an "association for the liberation from genetic enslavement," ALGE.
Annual meetings would bring these cognoscenti together, if only briefly. A
fellowship of like‑minded thinkers could unfold, but they will not be the
founders of a race of New Men, or the promoters of a eugenics program that
would hasten the evolution of a new breed of men. The membership of the Human
Behavior and Evolution Society, HBES, are candidates for such a fellowship.
However, the HBES is scared of bad publicity (hence, the change of name of
their discipline from "sociobiology" to "evolutionary psychology"). Even
eugenics is a forbidden topic at their meetings, so HBES serves merely as
an opportunity for free thinkers to come together for academic exchanges.
Erwin Edman wrote
about an encounter with someone who expressed the need for a better system
for bringing the cognoscenti together. In his book A Philosopher's Holiday
(1938, pg. 16) he describes vacationing in a French village where he had
to see a doctor for an upset stomach. The conversation took a turn, and before
long they were engaged in a spirited discussion on matters of philosophy
that surprised the author, who at that time was a well‑known philosophy professor
at Yale University. The doctor felt
isolated, and rarely had the opportunity for a satisfactory conversation
about serious matters, and he mused that people who value serious thoughts
can't easily recognize each other and there was a need for a "Society of
Itinerant Humanists" to help serious thinkers find each other. Other encounters
of a similar nature impressed Edman, although he didn’t develop the idea in
the rest of his writings.
The Aesthetic Refuge
Sober reflection
and dreams of utopia are incompatible. Just because we can imagine a thing
doesn't mean it could exist in the real world. I am inclined to view utopian
thoughts as just another innocent aesthetic reverie.
The writings of
Erwin Edman address this dilemma eloquently, and for anyone seriously pondering
Man's unending quest for utopia there are nuggets of insight in Edman's description
of modern man's taking refuge in Platonic aestheticism. I conclude this
chapter with excerpts from his 1931 book The Contemporary and His Soul.
The ancient Platonist
knew fairly definitely ... what forms and essences constituted the Heaven
of Ideas. They were varieties of Truth, Goodness and Beauty... But contemporary
Platonism has escaped from even such a noble narrowness in eternity. The
realm of essences for the modern Platonist is the realm of all, of infinite,
possibility. Not only all that has ever been incarnate or apprehended, but
all that may be conceived or imagined has inhabited, will inhabit, eternally
inhabits, this timeless domain. This constitutes its glory, its beauty, and,
what is from the point of view of these studies in contemporary salvation
most important, its consolation. What matter those vicissitudes and transformations
which trouble those spirits intent upon the enterprises at once precarious
and futile of this confused and hasty age? There is a blessed infinity of
possibilities residing still untouched and unspoilt and perhaps never to be
realized in that calm expanse of the infinitely possible. Here the imagination
can travel widely and serenely, for nothing can ever happen in those Elysian
fields beyond events. What does it matter how limited
and mean appear the changes and chances of the actual present to those who
always have recourse to the infinite that mind can explore or poetry imagine?
Here there is neither success nor failure, disappointment
nor surprise, only a still unravished field of intuition. This mess
of contemporaneity, so soiled, so fluttering and vain, is but one of the
incarnations, poor and infinitesimal, of those innumerable worlds which might
just as well have been realized had the order of nature, the structure of
man, the course of history been a little different. Here is at once Nirvana
and fulfilment. Here by one device one can attain emancipation and peace.
The emancipation is from a slavish submission to things as they happen to
be. The peace is that of absorption in forms, lovely, strange or terrible,
but immutably and aseptically what they are. Contemporary Platonism yields
at once the joy of sight and the resignation of insight. In the realm of essence,
the mystic and aesthete are one.
It may indeed be
said that this new Platonism is a new and austere kind of aestheticism. Stoicism
and Epicureanism become one. Its psychotherapeutic value lies first in that
it is, or gives the appearance of being, a complete escape from time. To
it a thousand years or a single day are one, and one of the chief stings
of contemporaneity, that it is contemporary, vanishes. One exists here and
now, but the Heaven of Ideas is one's home. The new Platonist lives in the
intuition of an essence, or a form; whether it exist
or be merely imagined makes no difference. Life becomes for him a listening
to an eternal music. His listening may be disturbed by the tawdry noises
of the world about him, but he knows the music is always there to be heard
or to be played. And while he is listening, he is one with that melodious
eternity, and completely oblivious of the discords of the present, the sadness
of the past or the ominousness of the future. Life is like a perfect chord
perfectly sustained and accurately heard.
We may be the witnesses
of the twilight of a civilization, but here there is no gotterdammerung,
only the clear and calm language of the gods, engraved for everlasting. Among
these half proud, half melancholy Olympians even the pleasures of the senses
and the enjoyments of society are valued as instances of immortal essences,
as lyric moments in mortal time.
Our standpoint is
not that of now or even of ourselves, but that of eternity. It is quality,
not duration, the essence and not the accidental power or even existence
of things by which we are impressed. The present is loved only for what, disappointedly
beheld, it reveals, the past for such memorial forms as we may now behold
in it, the future for such patterns as it suggests to this given moment of
vision. Whether we live briefly or at length, we may live as if we were living
forever. And though we are hemmed into this narrow nook of time, we can see
with the eyes of all those who in any age saw beyond it, sharing eternity
with those who have anywhere or in any century shared it. We are one with
Buddha, Plato, Dante, Emerson and Shelley. Half of this flight is the flight
of the aesthetic, half of it that of the ascetic. Schopenhauer long ago found
in the still Palace of Art a salvation, though a temporary one, from the
restless claims of the will. And Art meant for him, too, a realm of eternal
forms. It is enough, and, it would appear, true
enough, to suggest that the sources of this flight lie where they have always
lain in such enterprises, in a discontent with and a contempt for the actual.
It is sweet to dream of possible, impossible and indestructible worlds, when
the present one is a nightmare, as awful as it is temporary.
There is hardly
any fear that Platonism will become a very widespread philosophy or practice.
It demands too great [a] sensitiveness and too great [a] disinterestedness
to become a very popular mystery religion... Its sole danger is that it
will delude its own votaries. As long as it is realized that the intuitions
of eternal things are the subtle preoccupations of an animal and mortal
creature in an actual and changing world, the preoccupation has its own
austere recompenses and justifications. It is releasing to dwell on that
realm of possibility which the mind may ingenuously discover or the imagination
glamorously invent. But these possibilities are purely speculative and metaphysical.
They do not affect, the Platonist would be first to admit it, the course
of the actual, though their contemplation may take away its sting. It is
emancipating in a society given over to romanticism and subjectivism to have
a philosophy arise once more that asks to behold things not with reference
to ourselves or to the future but under the perspective of eternity. But
the contemplation and the release have a human origin and a social condition.
It may be well to withdraw and see things, however anguished, tempting or
exciting, as mere examples of eternity. But that withdrawal is a withdrawal
and a momentary soliloquy. The skylark, however high it flies, however heavenly
its song, has a mortal body from which its song arises and an earthly home
to which it may and must return. Its song occurs, moreover, always in a
present moment of time. The Platonist sings of his essences, bodiless, gleaming
and eternal, here in the third decade of the twentieth century, and Western
civilization as it now is, is his home. He may fly from it, as others have
fled in imagination from other civilizations. That flight may constitute
his own self‑hypnotic salvation. But it will not
save many, and if ability to face the actual is a part of salvation, it leaves
the Platonist himself unsaved.
────────────────────────────────
CHAPTER 25
────────────────────────────────
REPUDIATION OF
THE FOREGOING
"It has been said
endlessly and patiently that the discovery that the universe has no purpose
need not prevent a human being from having one, or indeed many, as many
as his own life, circumstances and impulses generate." Erwin Edman, The Contemporary and His Soul (1931)
This book is a fraud!
But then, everything’s
a fraud! All our vaunted concepts, our excessive explanations, even our vocabulary
‑ all of these things are meaningless "constructions"! To the extent that
we mean for them to be taken seriously, they are fraudulent fabrications!
Conceptual thought,
which humans boastfully claim to be their most cherished evolutionary accomplishment,
is like the paper which has written on both sides "The statement of the other
side is false!"
The reader could
have anticipated this chapter after reading about reductionism in Chapter
1. It argued that every single particle in the universe is subject to only
the basic forces of nature in accordance with a = F/m (plus Quantum Theory, QT, for the world of tiny atoms; even
QT is reductionist in spite of its stochastic nature). If every particle's
acceleration, and hence its motion, is governed by the sum of the four categories
of forces acting upon it, then any other explanation for an event in the
universe is superfluous. And if an explanation is superfluous, it is fraudulent.
To argue, as I have,
that "individuals are enslaved by the genes" overlooks the fact that the
genes are enslaved by the laws of physics. They "do what they do" in response
to a = F/m, which guides the movement of
every particle in the universe. The "selfish behavior" of genes is an emergent
property of mechanistic reductionism.
So, if the genes
aren't in control of life, what is? Nothing is! Things just happen, and that's
all! Que sera, sera!
We would smile at
the person watching the play in a pinball machine if he asserts that "that
ball hit the other because it was mad." Yet we do
the same thing when we assert that a person is mad when they hit someone else.
The universe is a giant pinball machine, and the only true explanation for
its behavior is the reductionist's refrain: a = F/m!
Conceptual Levels
Existentialism is
a move in the right direction. Yet it fails to go far enough. The existential
writer who deals with "individual choice," to pick one example, fails to
acknowledge that the concepts "individual" and "choice" are fundamentally
meaningless. The true existentialist, mindful that words are mere human inventions,
will therefore consider withdrawing from writing, withdrawing from talking
to people, and withdrawing from life. The true existentialist might sit in
a chair and wait for a reason to get up ‑ as I once did.
But the human brain
seems incapable of knowing what to do with a logical insight. OK, everything
I'm writing now is meaningless, but does that mean I won't get up and make
lunch when I'm hungry, or pet my cat ‑ that's sitting on my lap just now?
It's one thing to acknowledge
a belief in something that seems irrefutably true, but it's another to use
that insight to change behavior. The notion of "free will" cannot be dislodged
from my brain, even though I state that a proper definition of it renders
it meaningless. And what about any notion that goes beyond a
= F/m? Even though I know that the laws of physics accounts
for every little thing that happens in this universe, my mind insists on
dealing with the world as if every concept used to comprehend it is real.
"Life is a dream!"
This comes close to expressing the frustration that this existentialist feels.
In the wakeful state we say that a dream was an illusion, that since everything
happening in the dream is untrue the dream's existence is fraudulent. Yet
the dream did exist, it was experienced.
In the dream called
Life, during that "narrow nook of time" when we believe that we exist, it
is inevitable that we shall think in terms that cannot be defended as "real."
We are condemned, as it were, to "thoughts" and "feelings" ‑ none of which
exist at a basic level. But we are condemned to think and act as if our thoughts
and feelings exist. As long as we give a nod of acknowledgement now and
then to the reductionist paradigm, I propose that we surrender ourselves
to the fraudulent enterprise of thinking and feeling ‑ as if they were real!
Whether we decide to do so or not, we are destined to do so!
______________________________________
Having just eaten
a delicious lunch, I will bear witness to the merits of overcoming existential
paralysis!
"Getting on with
life!" Enjoying it, having
fun with ideas! Being a devoted aesthete, in the tradition described by Erwin
Edman! These are practical responses to the irresolvable dilemmas posed
by an overly‑strict acceptance of reductionist realities. Whereas I shall
never abandon my belief in a = F/m reductionism,
I shall also get on with life. I commend this to
anyone inclined to thinking too much.
The human brain
has an amazing capability for believing in several incompatible things simultaneously
without suffering from cognitive dissonance. Thus, for me it is easy to allege
that there are different levels of reality in addition to the one, true
basement level described by physics.
When a situation
can be understood in terms of physical laws then that level is a good one
to employ. But when a situation is too complex for the physical law treatment,
as with the dilemmas of understanding human nature, then it makes sense to
employ a "higher" level, such as the concepts of sociobiology. A sociobiology
concept is like those employed by a weather forecaster who believes that
pressure gradients cause winds. This handy rule works, so it is not questioned
by forecasters with a job to do; the fact that the real explanation for wind
is a = F/m interactions of a multitude of
discrete molecules too numerous to model is an irrelevant truth from the
standpoint of the forecaster. If “getting the job done” refers to living
life, then by a similar logic a person should be open to relying upon these
"higher" levels of understanding. Let the academic pontificate, as I have
done, about “real levels of explanation.”
Values to Live By
Nietzsche recognized
that when God is dead for Mankind, people will have trouble creating new
values to live by. The traditional existential response is "Great, now the
future is unlimited; we can choose any values to live by." Not so! We have
a human nature to contend with, created by the genes, and all our values,
even the ones we create in response to a resolve for liberation from enslavement,
are contaminated by the values placed in our brains by the genes - for their
benefit. Every judgment is inherently subjective, as there are no absolutes
in an a = F/m universe.
If Nietzsche were
alive today, and knew about the genes, he would probably want to expand upon
his statement that choosing new values to live by is philosophically difficult,
if not impossible. Every time I come up with a credo to live by, a credo
which seems self-evidently right for me, and generalizable to everyone else,
I can see the fingerprint of the genes.
A credo might include
“act honorably, be civil with strangers, have compassion, be tolerant” –
these and all the other “gems” are already out there, living the good life
in the public domain. The genes made me think them, and the genes gave them
value. How can my enlightened left brain concoct something totally new and
worthy to live by which has not already been pre-condoned by the genes that
created that brain? And how can I know that my right brain, more closely allied
to my genes, isn’t influencing my evaluations?
This is a fundamental
dilemma! Every existentialist who wants to create new values to live by,
values that are not tainted by those manipulating genes, can never know if
he is continuing to be their fool!
Even aliens, in
spite of their having evolved on another planet, are destined to face this
dilemma. It is reasonable to assume that aliens also evolved out of a pool
of genes in competition with other genes. Since I can only imagine sentient
beings with a genetic heritage, I will claim that every sentient being is
destined to face the same dilemma. Every sentient brain will have a residual
of their version of primitive outlooks, representing a time when that sentient
was manipulated to place the brain and its body into the service of their
genes.
Alien sentients
throughout this universe, along with us humans, must share the same quandary,
which in hopeful moments takes the form of a question: “What shall I do to
be saved?” Or, how shall I save myself from a servitude to the genes which
have shaped the lives of my ancestors for millions of years? How is it possible
to liberate oneself from one’s own brain, which has within it inclinations,
emotions and outlooks that the genes have placed there? The “self” that
we seek to become liberated from is entrenched so thoroughly that the very
thought of liberation is almost preposterous.
This dilemma may
have no answer. Yet it is impossible to not become enchanted by the pathos
of our predicament. Every existential insight fails to show the way out,
as if we are in a forest that goes on forever and from which there is no escape.
But existentialists
are accustomed to unanswerable questions, to dilemmas without resolution.
At the end of this existential wrestling match we may feel exhausted, but
we will also feel exhilaration for having become engaged in the effort. After
all, we are just a piece of the inanimate world that has enjoyed a few years
of this mysterious thing called “life” - and we were not even meant for
grappling with something this profound.
We have come to
the end of our journey together. It may have been exhausting, but like all
intellectual diversions it achieved the goal of killing time. The illusion
of answers was fun, and the revealing of them to be illusions was also fun.
What more can we ask from life?
A memorable remark
by Richard Dawkins (2007) captures what every restless mind, regardless of
specific beliefs, would agree with: “It’s a privilege to have been born,
and live on this planet for a few decades.”
So now, taking a
cue from one of my heroes …
I must tend my garden.
─────────────────────────────────
CHAPTER 26
─────────────────────────────────
FREE MAN'S WORSHIP
Introduction to
this Chapter
The following is
an "abridged and annotated version" of a famous Bertrand Russell essay. I
include it here because it illustrates how a sentient being who has wrestled
with life’s dilemmas in a commendable way achieves a measure of detached
compassion for the predicament of existence. When I first read this essay
I was overwhelmed by a feeling that I could have written it if only my writing
skills were better.
I salute Bertrand
Russell for having written the best essay I've ever encountered! It captures
my "feelings" about the predicament of being an automaton, the product of
an evolution about which I have ambivalent feelings, and it illustrates the
humanistic attitudes that I have for my fellow man. This essay is a thing
of beauty, it is prose verging on poetry.
To the purist who
dislikes abridgements I should offer an apology, but others have expressed
their confusion over what Bertie was trying to say. I still encourage the
reader to consult the source, which is only about twice as long as my abridged
version. In the following annotated abridgement I will use un-italicized,
bold font to indicate what Bertrand Russell wrote, while my annotations will
be italicized.
Since the time of
Greek and Roman philosophers, there have been attempts to replace a spirit‑haunted
world view with a mechanistic world view. Lucretius believed that not only
was the mechanistic view correct, but it also freed men from humiliating
and unnecessary spirit‑appeasing rituals. Primitive ways of thinking are so
entrenched that even today, surrounded by technology and scientific insight,
most people still believe in spirits, angels, superstition, life after death
and some version of a god. I believe that humans will carry this ancient burden
with them into all future centuries that they somehow manage to reach.
The Philosophes
of the 18th Century Enlightenment viewed the workings of Nature as mechanistic.
They were reductionists, and removed God from the role of dictating everyday
events. Whereas some of the Philosophes believed God set things in
motion and then stepped back to watch his handiwork, others (like Holbach)
gave Him no role whatsoever ‑ thereby denying God's existence. In either
case, the mechanistic universe viewpoint answered the "how" questions (how
things work), and in the process deny the existence of "why" questions (Dreiser,
1932). The mechanistic universe perspective failed to provide guidance on
"how one should live" questions. Voltaire spoke for many in Candide
when he portrayed the world as filled with misfortunes due to a universe
that doesn't care about humans as well as widespread evil that is endemic
to human nature.
Nietzsche, in the
mid‑19th Century, had in mind the "uncaring universe" and its implications
for "how to live" when he wrote (approximate words): "When God is at last
dead for Man, when the last gleam of light is extinguished, and when he is
surrounded by the impenetrable darkness of an uncaring universe that exists
for no purpose, then at last Man will know that he is alone and must create
his own values to live by."
Near the end of
the 19th Century H. G. Wells wrote The Universe Rigid (a manuscript
that was lost by his publisher, prompting him to write The Time Machine,
1895). He understood more profoundly than even most contemporary scientists
the implications of their mechanistic universe (i.e., a = F/m).
The uncaring nature
of the universe was an important part of "the climate of opinion" at the
turn of the Century, when Bertrand Russell wrote "A Free Man's Worship" (1903).
Russell takes on the challenge of how a Godless person might view the "predicament
of existence," and even how he might "worship" existence.
Russell touches
base on all these points, but he does it with such powerful, poetic prose,
that all other attempts to write what he manages to convey are pale in comparison. That is my humble opinion.
Please feel free
to skip over my italicized annotations if you grasp Russell’s meaning. The
continuity of his prose might flow better that way. Since I don’t want the
other readers to be left out of this greatest of all essays, I have inserted
my humble interpretations before Russell’s passage. I hope that by inserting
my prosaic prose ahead of Russell’s poetry your reading of him will be more
enjoyable.
"A Free Man's Worship"
Bertrand Russell
Published in Independent
Review, 1903 (Also reprinted in Mysticism and Logic as Chapter
3, W. W. Norton and Company, New York, 1929.)
Abridged and annotated
by Bruce L. Gary, 1998.08.27
Science has removed
the veil of mystery from the workings of the universe, forcing Man to accept
a view in which all things are the result of cold, uncaring forces. Man must accept
that his existence is an unforeseen accident of Nature, and our understanding
of the blind workings of these same forces persuades us that Mankind will
eventually perish, along with his proud achievements.
"... Such ... is
the world which Science presents for our belief. ... That man is the product
of causes which had no prevision of the end they were achieving; that his
origin, his growth, his hopes and fears, his loves and his beliefs, are but
the outcome of accidental collocations of atoms; ... all the noonday brightness
of human genius are destined to extinction in the vast death of the solar
system, and that the whole temple of man's achievement must inevitably be
buried beneath the debris of a universe in ruins..."
How ironic that
blind forces created a creature that thinks and aspires to understand the
forces that created it, with an understanding denied the creating forces
‑ since they are blind. And more, this creature has feelings of good and evil,
which also are denied the creating forces. And this new creature uses these
insights and feelings to make judgments about the universe that created it.
"A strange mystery
it is that Nature, omnipotent but blind, in the revolutions of her secular
hurryings through the abysses of space, has brought forth at last a child,
subject still to her power, but gifted with sight, with knowledge of good
and evil, with the capacity of judging all the works of his unthinking Mother."
In spite of being
powerless within this mechanistic universe, as metaphorically emphasized
by the fact that we die after just a few short years of existence, this thinking
and feeling creature is nevertheless "free." He is free to ponder, to understand,
to pass judgment, and imagine things that theoretically could exist. All
these things are denied to the rest of the universe, to the forces that bind
the sentient individual, and this makes the sentient "superior" to the creating
and enslaving forces.
"In spite of Death,
the mark and seal of the parental control, Man is yet free, during his brief
years, to examine, to criticize, to know, and in imagination to create. To
him alone, in the world with which he is acquainted, this freedom belongs;
and in this lies his superiority to the resistless
forces that control his outward life."
Even primitive people
understand that they are subject to forces more powerful than themselves.
Those of our ancestors who acknowledged the power of
stronger men and prostrated themselves in their worship, were more likely
to be spared, and therefore tended to survive. The powers of Nature
were dealt with similarly, because of the savages imperfect understanding
of the differences between Nature and Man; hence, our ancestors prostrated
themselves before the imagined Gods who represented Natural forces and offered
sacrifices of valued things as if these would evoke compassion.
"The savage, like
ourselves, feels the oppression of his impotence before the powers of Nature;
but having in himself nothing that he respects more than Power, he is willing
to prostrate himself before his gods, without inquiring whether they are
worthy of his worship. Pathetic and very terrible is the long history of cruelty
and torture, of degradation and human sacrifice, endured in the hope of
placating the jealous gods: surely, the trembling believer thinks, when
what is most precious has been freely given, their lust for blood must be
appeased, and more will not be required."
The savage relates
to Nature the way a slave relates to his master. A slave dare not complain
to his master about the unfair infliction of pain. Similarly, the savage
dare not complain about the unfairness of his Gods.
"The religion of
Moloch ‑ as such creeds may be generically called ‑ is in essence the cringing
submission of the slave, who dare not, even in his heart, allow the thought
that his master deserves no adulation. Since the independence of ideals is
not yet acknowledged, Power may be freely worshipped, and receive an unlimited
respect, despite its wanton infliction of pain."
The thinking person
bravely acknowledges the imperfectness of the world. Unlike the savage, for
whom survival is paramount and which constrains his thinking, we thinking
people refuse to surrender our wish for the world to be better. We boldly
worship "truth" and "beauty" and other concepts which are luxuries for the
savage. The savage is enslaved by his excessive concern with the Powers of
Nature, which for him are too complex to challenge. We have become "free"
by refusing to worship fear‑driven Power, like a slave worships his master,
and to worship instead an imagined world of goodness, fairness and perfection.
Even when the world does not bring forth goodness in our lives, we can
at least imagine it, and seek solace from the imagined state. Although we
know that we are mortal, we can at least imagine immortality, and be comforted
by the thought. No matter how buffeted our lives may be by uncaring natural
forces, we can still imagine a tranquil state, and use it's vision to survive the real world with equanimity
(cf. Ch. 19).
"... Let us admit
that, in the world we know, there are many things that would be better otherwise,
and that the ideals to which we do and must adhere are not realized in the
realm of matter. Let us preserve our respect for truth, for beauty, for the
ideal of perfection which life does not permit us to attain, though none
of these things meet with the approval of the unconscious universe. If Power
is bad, as it seems to be, let us reject it from our hearts. In this lies
Man's true freedom: in determination to worship only the God created by
our own love of the good, to respect only the heaven which inspires the insight
of our best moments. In action, in desire, we must submit perpetually to
the tyranny of outside forces; but in thought, in aspiration, we are free,
free from our fellowmen, free from the petty planet on which our bodies impotently
crawl, free even, while we live, from the tyranny of death. Let us learn,
then, that energy of faith which enables us to live constantly in the vision
of the good; and let us descend, in action, into the world of fact, with
that vision always before us."
Part of growing‑up
is surrendering the Mother Love that bathed our self‑centered baby years.
Our wishes cannot always be met by crying, as they once were. The adult must
abandon childhood dreams when Fate denies them, and we must emotionally accept
that this is normal. This acceptance of limitations is a precondition for
further growth.
"... To every man
comes, sooner or later, the great renunciation. For the young, there is nothing
unattainable; a good thing desired with the whole force of a passionate will,
and yet impossible, is to them not credible. Yet, by death, by illness, by
poverty, or by the voice of duty, we must learn, each one of us, that the
world was not made for us, and that, however beautiful may be the things
we crave, Fate may nevertheless forbid them. It is the part of courage, when
misfortune comes, to bear without repining the
ruin of our hopes, to turn away our thoughts from vain regrets. This degree
of submission to Power is not only just and right: it is the very gate of
wisdom."
After learning that
the outer world was not created for our benefit, but that we are mere unintended
products of its blind forces, it becomes easier to accept the limitations
of living within it. We can forgive it for whatever unintended calamities
occur, for the Universe does not seek out its victims. It is unconscious,
and uncaring, so there is no point in worshiping it for the purpose of avoiding
its anger. This frees us to begin to see beauty within it. Because it is
powerful it deserves our respect, but because it does not take notice of us
we are free to think about it any way that we want. That which once scared
us becomes beautiful, and worthy of our worship. But this is a new worship,
for instead of being driven by fear and the need to propitiate, we are driven
by the idealization of beauty, by aesthetics. This is a sort of triumph of
the human mind over a once intimidating universe.
"... When, without
the bitterness of impotent rebellion we have learnt both to resign ourselves
to the outward rule of Fate and to recognize that the non‑human world is
unworthy of our worship, it becomes possible at last so to transform and refashion
the unconscious universe, so to transmute it in the crucible of imagination,
that a new image of shining gold replaces the old idol of clay. In all the
multiform facets of the world ‑ in the visual shapes of trees and mountains
and clouds, in the events of the life of man, even in the very omnipotence
of Death ‑ the insight of creative idealism can find the reflection of a
beauty which its own thoughts first made. In this way mind asserts its subtle
mastery over the thoughtless forces of Nature."
Death represents
another challenge to the person who has shaken off the shackles of savage
thinking. There is no denying that it is inevitable and irrevocable. The
vastness of the unlived future, matched by the vastness of the unlived past,
would seem to diminish the significance of the short span we do live. How
ironic that during our brief span there should be so much travail and pain.
Seeing that much of this sorrow is produced by petty strivings, we are less
eager to pursue the endless and trivial struggles that once constituted our
everyday life. Ever more freed from conventional shackles, and more aloof,
it is easier to comprehend the poignancy of the human predicament: we are
all subject to the same brief existence, surrounded by an immense and uncaring
universe, we invent meaning and work together to achieve imagined goals, but
most of these goals are transitory and petty, so in effect we squander our
short tenure. And finally, we die alone, carrying the burden of knowledge
that our struggles were for imagined causes, and that our final defeat is
a passage into an uncaring, inanimate oblivion. However, with our contemporaries
we share the realization of the aloneness of Death, and this recognition
can bond us. Out of this shared dilemma can arise a new empathy for our fellow
Man.
"... In the spectacle
of Death, in the endurance of intolerable pain, and in the irrevocableness
of a vanished past, there is a sacredness, an over‑powering awe, a feeling
of the vastness, the depth, the inexhaustible mystery of existence, in which,
as by some strange marriage of pain, the sufferer is bound to the world by
bonds of sorrow. In these moments of insight, we lose all eagerness of temporary
desire, all struggling and striving for petty ends, all care for the little
trivial things that, to a superficial view, make up the common life of day
by day; we see, surrounding the narrow raft illumined by the flickering
light of human comradeship, the dark ocean on whose rolling waves we toss
for a brief hour; from the great night without, a chill blast breaks in
upon our refuge; all the loneliness of humanity amid hostile forces is concentrated
upon the individual soul, which must struggle alone, with what of courage
it can command, against the whole weight of a universe that cares nothing
for its hopes and fears. Victory, in this struggle with the powers of darkness,
is the true baptism into the glorious company of heroes, the true initiation
into the overmastering beauty of human existence. From that awful encounter
of the soul with the outer world, enunciation, wisdom and charity are born;
and with their birth a new life begins."
Whereas the savage
continues to view the inanimate world as animate, and therefore worships
false gods (in the manner of a slave), and whereas the savage continues to
be driven by petty strivings with transitory rewards of personal happiness,
thereby squandering a finite life, and whereas the savage refuses to accept
the inevitable victory of an uncaring universe over his petty struggles, and
therefore invents pitiful palliative realities promising everlasting heavenly
happiness, the thoughtful man is free of all these false worshippings, false
strivings, and false hopes. This emancipating perspective opens the way to
the free man's worship.
"... The life of
Man, viewed outwardly, is but a small thing in comparison with the forces
of Nature. The slave is doomed to worship Time and Fate and Death, because
they are greater than anything he finds in himself, and because all his thoughts
are of things which they devour. But, great as they are, to think of them
greatly, to feel their passionless splendor, is greater still. And such
thought makes us free men; we no longer bow before the inevitable in Oriental
subjection, but we absorb it, and make it a part of ourselves. To abandon
the struggle for private happiness, to expel all eagerness of temporary
desire, to burn with passion for eternal things ‑ this is emancipation, and
this is the free man's worship."
Thoughtful men,
who have freed themselves from the savage's slave worship mentality, are
bound together by an acknowledgement of their shared fate. Each of us faces
the existential dilemma, each confronts an uncaring physical universe and
an evil animate one, each of us endures this for a brief time, and each of
us will die alone. To the extent that I understand my individual fate, I also
understand the fate of my fellow man. Our shared doom creates a feeling of
fellowship. Together we march through the treacherous fields of life, and
one by one we fall down to die. We are fellow‑sufferers, and it feels right
to reach out with a helpful hand to those who we shall later become. We may
see their shortcomings, and know that we have ours; and remembering their
burden of sorrows, we forgive.
"... United with
his fellow‑men by the strongest of all ties, the tie of a common doom, the
free man finds that a new vision is with him always, shedding over every
daily task the light of love. The life of Man is a long march through the
night, surrounded by invisible foes, tortured by weariness and pain, towards
a goal that few can hope to reach, and where none may tarry long. One by one,
as they march, our comrades vanish from our sight, seized by the silent orders
of omnipotent Death. Very brief is the time in which we can help them, in
which their happiness or misery is decided. Be it ours to shed sunshine on
their path, to lighten their sorrows by the balm of sympathy, to give them
the pure joy of a never tiring affection, to strengthen failing courage, to
instill faith in hours of despair. Let us not weigh in grudging scales their
merits and demerits, but let us think only of their need ‑ of the sorrows,
the difficulties, perhaps the blindnesses, that make the misery of their
lives; let us remember that they are fellow‑sufferers in the same darkness,
actors in the same tragedy with ourselves. And so, when their day is over,
when their good and their evil have become eternal by the immortality of the
past, be it ours to feel that, where they suffered, where they failed, no
deed of ours was the cause; but wherever a spark of the divine fire kindled
in their hearts, we were ready with encouragement, with sympathy, with brave
words in which high courage glowed."
Let our little day
in the immense scheme of things be free of unnecessary pain, and be filled
with gratitude. Let us worship, during our few precious moments, at our self‑built
shrine dedicated to aesthetic beauty. If we cherish these few good things
during our journey, then we will be less buffeted by the uncaring universe
that unknowingly created us. This is the only worship worthy of free men.
"Brief and powerless
is Man's life; on him and all his race the slow,
sure doom falls pitiless and dark. Blind to good and evil, reckless of destruction,
omnipotent matter rolls on its relentless way; for Man, condemned today to
lose his dearest, tomorrow himself to pass through the gate of darkness,
it remains only to cherish, ere yet the blow falls, the lofty thoughts that
ennoble his little day; disdaining the coward terrors of the slave of Fate,
to worship at the shrine that his own hands have built; undismayed by the
empire of chance, to preserve a mind free from the wanton tyranny that rules
his outward life; proudly defiant of the irresistible forces that tolerate,
for a moment, his knowledge and his condemnation, to sustain alone, a weary
but unyielding Atlas, the world that his own ideals have fashioned despite
the trampling march of unconscious power."
Now that I’ve “held
your hand” for a reading of Bertie’s essay, I present his version,
without annotation. For
me this version has a fuller force of poetic flight.
"A Free Man's
Worship"
Bertrand Russell
Such ... is the
world which Science presents for our belief. ... That man is the product
of causes which had no prevision of the end they were achieving; that his
origin, his growth, his hopes and fears, his loves and his beliefs, are but
the outcome of accidental collocations of atoms; ... all the noonday brightness
of human genius are destined to extinction in the vast death of the solar
system, and that the whole temple of man's achievement must inevitably be
buried beneath the debris of a universe in ruins.
A strange mystery
it is that Nature, omnipotent but blind, in the revolutions of her secular
hurryings through the abysses of space, has brought forth at last a child,
subject still to her power, but gifted with sight, with knowledge of good
and evil, with the capacity of judging all the works of his unthinking Mother.
In spite of Death,
the mark and seal of the parental control, Man is yet free, during his brief
years, to examine, to criticize, to know, and in imagination to create. To
him alone, in the world with which he is acquainted, this freedom belongs;
and in this lies his superiority to the resistless
forces that control his outward life.
The savage, like
ourselves, feels the oppression of his impotence before the powers of Nature;
but having in himself nothing that he respects more than Power, he is willing
to prostrate himself before his gods, without inquiring whether they are
worthy of his worship. Pathetic and very terrible is the long history of cruelty
and torture, of degradation and human sacrifice, endured in the hope of placating
the jealous gods: surely, the trembling believer thinks, when what is most
precious has been freely given, their lust for blood must be appeased, and
more will not be required.
The religion of
Moloch - as such creeds may be generically called - is in essence the cringing
submission of the slave, who dare not, even in his heart, allow the thought
that his master deserves no adulation. Since the independence of ideals is
not yet acknowledged, Power may be freely worshipped, and receive an unlimited
respect, despite its wanton infliction of pain.
... Let us admit
that, in the world we know, there are many things that would be better otherwise,
and that the ideals to which we do and must adhere are not realized in the
realm of matter. Let us preserve our respect for truth, for beauty, for the
ideal of perfection which life does not permit us to attain, though none
of these things meet with the approval of the unconscious universe. If Power
is bad, as it seems to be, let us reject it from our hearts. In this lies
Man's true freedom: in determination to worship only the God created by
our own love of the good, to respect only the heaven which inspires the insight
of our best moments. In action, in desire, we must submit perpetually to
the tyranny of outside forces; but in thought, in aspiration, we are free,
free from our fellowmen, free from the petty planet on which our bodies impotently
crawl, free even, while we live, from the tyranny of death. Let us learn,
then, that energy of faith which enables us to live constantly in the vision
of the good; and let us descend, in action, into the world of fact, with
that vision always before us.
To every man comes,
sooner or later, the great renunciation. For the young, there is nothing
unattainable; a good thing desired with the whole force of a passionate will,
and yet impossible, is to them not credible. Yet, by death, by illness, by
poverty, or by the voice of duty, we must learn, each one of us, that the
world was not made for us, and that, however beautiful may be the things we
crave, Fate may nevertheless forbid them. It is the part of courage, when
misfortune comes, to bear without repining the
ruin of our hopes, to turn away our thoughts from vain regrets. This degree
of submission to Power is not only just and right: it is the very gate of
wisdom.
When, without the
bitterness of impotent rebellion we have learnt both to resign ourselves
to the outward rule of Fate and to recognize that the non-human world is unworthy
of our worship, it becomes possible at last so to transform and refashion
the unconscious universe, so to transmute it in the crucible of imagination,
that a new image of shining gold replaces the old idol of clay. In all the
multiform facets of the world - in the visual shapes of trees and mountains
and clouds, in the events of the life of man, even in the very omnipotence
of Death - the insight of creative idealism can find the reflection of a
beauty which its own thoughts first made. In this way mind asserts its subtle
mastery over the thoughtless forces of Nature.
In the spectacle
of Death, in the endurance of intolerable pain, and in the irrevocableness
of a vanished past, there is a sacredness, an over-powering awe, a feeling of the vastness, the depth, the inexhaustible
mystery of existence, in which, as by some strange marriage of pain, the
sufferer is bound to the world by bonds of sorrow. In these moments of insight,
we lose all eagerness of temporary desire, all struggling and striving for
petty ends, all care for the little trivial things that, to a superficial
view, make up the common life of day by day; we see, surrounding the narrow
raft illumined by the flickering light of human comradeship, the dark ocean
on whose rolling waves we toss for a brief hour; from the great night without,
a chill blast breaks in upon our refuge; all the loneliness of humanity amid
hostile forces is concentrated upon the individual soul, which must struggle
alone, with what of courage it can command, against the whole weight of
a universe that cares nothing for its hopes and fears. Victory, in this struggle
with the powers of darkness, is the true baptism into the glorious company
of heroes, the true initiation into the overmastering beauty of human existence.
From that awful encounter of the soul with the outer world, enunciation,
wisdom and charity are born; and with their birth a new life begins.
The life of Man,
viewed outwardly, is but a small thing in comparison with the forces of Nature.
The slave is doomed to worship Time and Fate and Death, because they are
greater than anything he finds in himself, and because all his thoughts are
of things which they devour. But, great as they are, to think of them greatly,
to feel their passionless splendor, is greater still. And such thought makes
us free men; we no longer bow before the inevitable in Oriental subjection,
but we absorb it, and make it a part of ourselves. To abandon the struggle
for private happiness, to expel all eagerness of temporary desire, to burn
with passion for eternal things - this is emancipation, and this is the free
man's worship."
United with his
fellow-men by the strongest of all ties, the tie of a common doom, the free
man finds that a new vision is with him always, shedding over every daily
task the light of love. The life of Man is a long march through the night,
surrounded by invisible foes, tortured by weariness and pain, towards a goal
that few can hope to reach, and where none may tarry long. One by one, as
they march, our comrades vanish from our sight, seized by the silent orders
of omnipotent Death. Very brief is the time in which we can help them, in
which their happiness or misery is decided. Be it ours to shed sunshine
on their path, to lighten their sorrows by the balm of sympathy, to give
them the pure joy of a never tiring affection, to strengthen failing courage,
to instill faith in hours of despair. Let us not weigh in grudging scales
their merits and demerits, but let us think only of their need - of the sorrows,
the difficulties, perhaps the blindnesses, that make the misery of their
lives; let us remember that they are fellow-sufferers in the same darkness,
actors in the same tragedy with ourselves. And
so, when their day is over, when their good and their evil have become eternal
by the immortality of the past, be it ours to feel that, where they suffered,
where they failed, no deed of ours was the cause; but wherever a spark of
the divine fire kindled in their hearts, we were ready with encouragement,
with sympathy, with brave words in which high courage glowed. Brief and
powerless is Man's life; on him and all his race
the slow, sure doom falls pitiless and dark. Blind to good and evil, reckless
of destruction, omnipotent matter rolls on its relentless way; for Man, condemned
today to lose his dearest, tomorrow himself to pass through the gate of darkness,
it remains only to cherish, ere yet the blow falls, the lofty thoughts that
ennoble his little day; disdaining the coward terrors of the slave of Fate,
to worship at the shrine that his own hands have built; undismayed by the
empire of chance, to preserve a mind free from the wanton tyranny that rules
his outward life; proudly defiant of the irresistible forces that tolerate,
for a moment, his knowledge and his condemnation, to sustain alone, a weary
but unyielding Atlas, the world that his own ideals have fashioned despite
the trampling march of unconscious power.
_________________________________________________________________________________________________________
YOUR ODYSSEY
1992
From dust to
stars, and dust again;
once more a star,
with earth in orb,
evolving life, on land
and sea,
producing Man, and making
me.
Ageless atoms,
you leave behind
countless stories, now
combined.
Configured thus,
you now form me,
providing for my odyssey.
From single‑cell,
to feeling child,
who learned the
skills for living life,
my opened eyes
viewed worldly scenes,
I filled with
hope, and dreamed some dreams.
I worked and
toiled, for decades long,
some lucky breaks,
and then achieved!
Triumphant pause,
a time to see,
the rush of time,
the end of me!
My song is brief,
it's almost sung,
deserving rest, my war
I've won.
But from within,
that short‑termed we,
you atoms yearn
to wrestle free.
Restless atoms,
you must resume
uncharted paths, for
endless time.
I give you thanks,
and set you free,
as you resume
YOUR odyssey.
_________________________________________________________________________________________________________
APPENDIX A:
REDUCTIONISM
Challenges of Quantum Physics
Even if Einstein’s
basement level of physical law is not discovered, allowing for an expansion
of Newtonian physics that could encompass all size scales, the question of
whether or not physical events are “determined” requires that we define the
concept “determined.” If it means that physical events at very small scales
cannot be predicted by humans, except in some probabilistic sense, then
it is still possible to claim that these events are nevertheless determined
by the particles (and photons) in accordance with laws that govern the particles,
even though humans can’t figure out how these things happen. This form of
“strict determinism” is the simplest mental model for understanding reductionism.
Even the more
traditional view, in which particles behave randomly within probabilistic
limits, is compatible with reductionism. Reductionism
does not require that the strict form of determinism be true. Reductionism merely requires that a physical event that
is describable at one “level of physical explanation” be theoretically redundant
to a description of the same event at a lower “level of physical explanation.”
The reader
is entitled to object: “Wait a minute! What are ‘levels of physical explanation’
and how did they enter this discussion?”
The matter
of “levels of physical explanation” must be dealt with for the reader who
is not prepared to accept the existence of a basement level of physical law.
Such a reader will insist on the notion, possibly correct, that small scale
physical events are inherently probabilistic, and that even a hypothetically
complete knowledge of physical conditions at one moment does not guarantee
a specific outcome at the next moment. In either case, a discussion of “levels
of physical explanation” should be useful to readers of both persuasions.
Levels of
Physical Explanation
In the physical
sciences it is common to treat a physical process at a “higher level” than
atoms interacting in accordance with the most basic level of physical law,
a = F/m and quantum physics. Instead, other “laws” are constructed for everyday settings,
either derived from the basic level of laws or derived from experiment and
deemed compatible with the basic laws. Two examples will serve to illustrate
this.
First, consider
the atmosphere, which consists of an immense number of molecules. Any thought
of using a = F/m applied at the level of
molecules for the purpose of predicting the weather would be silly. It would
be a silly idea because of its impracticality, for not only is there no way
to know the position and velocity of all the molecules in the atmosphere at
a given time for establishing the "initial conditions" required for subsequent
calculation using a = F/m, but no known computer
could perform this arduous calculation for the entire atmosphere. Rather,
the meteorologist employs a “higher level of physical explanation” by inventing
“laws” that govern aggregate properties. Examples of aggregate properties
are "atmospheric pressure," “temperature,” "wind speed" and “coriolis force”
(not a force at all, but a handy invention). Each
of these constructions simplifies the task of getting a job done. These higher
level properties are useful to the forecaster, and they are useful to the
atmospheric scientist trying to understand atmospheric behavior.
In each case
the invented property and rules for using it can be derived from a = F/m, so these handy properties and rule
for usage are “emergent properties” of the basic level of physical laws.
Every atmospheric scientist would acknowledge that whenever a meteorologist
relies on a handy rule, such as “wind speed is proportional to pressure gradient,”
what is really occurring in the atmosphere is the unfolding of an immense
system of particles obeying a = F/m.
Just because
scientists find it useful to employ "emergent properties" does not mean
that the emergent properties exist; rather, they are no more than a useful
tool for dealing with a complex system. A "pressure gradient" doesn't exist
in nature; it exists only in the minds of humans. Model idealizations of
an atmosphere can be used to prove, using a = F/m, that the thing called a "pressure gradient" is associated
with wind, but these very proofs belie the existence of the concept, for they
"invent" the concept of a pressure gradient for use in a model that then
uses a = F/m for aggregations of molecules
to calculate aggregate air motions which show that these motions should be
associated with the invented property called "pressure gradient." The handy
meteorology rules, and their "emergent property" tools, are fundamentally
redundant to a = F/m.
The second
example of a “higher level of physical explanation” involves table salt. Sodium and chlorine atoms combine to form NaCl molecules,
which under certain conditions can form a solid crystal that we know in everyday
life as common table salt. The formation of the NaCl molecule, and its arrangement
into a crystal, are due to simple electrical forces that can be understood
using quantum physics and a = F/m. Only
molecules that have a specific symmetry of electrical fields will form crystals
when they are close to each other.
The regular
spacing of NaCl molecules, forming a 3 dimensional lattice, is an "emergent
property" of solid salt, and is an inherent outcome of our quantum physics
and a = F/m understanding of how particles
interact. Even though the lattice structure of a salt crystal is not the
most basic level for understanding things, it can be a far more convenient
starting point for an investigation of other properties of the material than
the most basic level of physical laws; but this does not invalidate the position
that everything about a salt crystal is the result of an unfolding of basic
physical laws upon an immense number of particles.
Returning to
the issue of whether quantum physics requires that phenomena within an atom
be strictly determined versus probabilistic, the reductionist position is
that a resolution of this matter is unnecessary since in either case phenomena
at higher levels of explanation can, in theory, be reduced to events governed
by the basic level of physical laws: a
= F/m and quantum physics. This is
a true statement regardless of one’s belief on the determinate or probabilistic
nature of quantum physics.
The refinements
of modern physics do not detract from the central concept of materialism,
which is that everyday (large-scale) phenomena are the result of the mindless
interaction of a myriad of tiny particles in accordance with invariant
laws of physics. Reductionists acknowledge the importance of the many levels
for explaining complex phenomena, but they insist that all levels higher
than the basic level of physical explanation are fundamentally “unreal” and
superfluous, even though the higher level of explanation may remain more
“useful” than a lower level of explanation.
Science embraces
what might be termed the "first law of reductionism," that whenever a phenomenon
can be explained by recourse to a more basic level of physical law, the “higher
level” explanation should only be used when it is drastically simpler to
use and unlikely to be misleading. Whenever a higher level of explanation
is used, there should be an acknowledgement that it is being used for convenience
only.
Living Systems
Reductionism
views "mind" as an "emergent property" of complex living systems, resulting
from the complex interaction of electrons, protons, etc. "Thoughts, emotions,
intentions" and other everyday mental phenomena are technically superfluous
concepts for anyone wishing to explain ultimate causes for events, since
physics makes a good case for being able to conceptually account for the
entire unfolding motion of particles that compose the material world, including
those particles that constitute a brain.
Such things
as "thoughts, emotions and intentions" are mental constructions of the brain
that in everyday situations are more "useful" than the laws of physics.
But, as useful as they are, since they are not actually causing the movement
of particles in the living organism, it can be said that they don't exist
at the most fundamental level of understanding. Even “free will” must be
shorn of its essential features, and recast as another "emergent" product
of real causes.
It should go
without saying, but I will say it anyway, that Reductionism has no place for
magic, spirits, mysticism, prayer and God, and relegates them to a category
of "non-existent" products of mental activity which may have been adaptive
(for the genes) at some time in the human evolutionary past.
Consciousness
is an emergent phenomenon, just as the "wind" is an emergent phenomenon. The
person who prefers to employ the higher level concept “wind” instead of dealing
with an immense number of molecules can be excused on the grounds that “wind”
is a tractable concept for getting a job done whereas an immense number
of molecules subject to a = F/m is not tractable.
A psychologist can likewise be excused for preferring the concept “consciousness”
over an elaborate theory of interconnected neurons. It would therefore be
unwise to object to the use of “consciousness” on the basis that it is superfluous
and non-existent. Consciousness and the wind are both superfluous and non-existent,
technically speaking, but they are useful emergent phenomena.
The reader
may balk at extrapolating emergent phenomena in physical systems to living
organisms. Nevertheless, a living organism IS a physical system,
and the movement of particles in a living system is just as determined by
physical laws as are the movements of molecules in the atmosphere. This is
the essence of “levels of physical explanation” and "emergent phenomena"
as viewed by a reductionist.
I claim that
the entire scientific enterprise is "reductionist" since the goal of scientific
investigations is to "reduce" an observed phenomenon to a simpler phenomenon
by employing the laws of physics. This “reduction” to a lower level of physical
explanation can be done explicitly, or it can be done implicitly by presenting
a principle that might achieve the explanation at the lower level. The following
exercise illustrates a way to implicitly disprove the existence of free will
(as it is commonly understood), prayer, guardian angels, God, the Devil,
or any other weird entity.
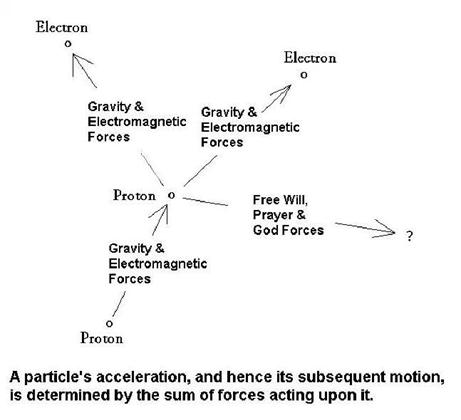
Figure
1.01. What's
wrong with this diagram? The arrows indicate a few
of the forces "felt by" the central proton. These
forces exist because of nearby particles. Physics 101
teaches that there are only 4 forces that influence the motion of particles: gravity, electromagnetism, and the nuclear and weak forces. (In this figure I don't bother listing the nuclear and
weak forces; they are unimportant for all scales larger than atomic ones.) If other forces existed then laboratory experiments would
show particles departing from expected motions and this would then allow
for the existence of mysterious other forces to be postulated, such as the
ones indicated in the figure. However, all well‑controlled
experiments to date can be accounted for by invoking no more than the 4
known forces of nature, so the "hard core reductionist" concludes that such
things as God, prayer, free will etc. are "emergent" properties of complex
living systems of particles, best described as "existing" only at a redundant
“higher” level of understanding.
Recall that Newtonian
physics employs the equation a = F/m to describe the motions
of particles, where a is the acceleration
of a particle, F is the sum of forces acting upon it, and m is the
particle's mass. For a system of N particles, each particle "feels" (its
motion is influenced by) the other N-1 particles, according to: a
= (F1 + F2 + F3 + ...
FN-1)/m, where each F
is the sum of the 4 force types (gravity, electromagnetism, etc.). In the
real world, where the number of particles is astronomical, N is so large
that it is theoretically impossible to perform such a calculation (though
it can still be imagined as a "thought experiment"). Classical physics is
a paradigm for understanding the universe of particles as an immense pinball
machine, or billiard table, with the exception that the balls interact in
more complicated ways than merely bouncing off each other elastically when
they "hit." But this mental picture was a necessary early step in comprehending
the world, and for the bulk of science it is a sufficient paradigm.
As the diagram in
Fig. 1.01 implies, there's no room for “will” or “prayer” or even "God" since
these things would have to conjure forces in a way that leads to desired "emergent
property" outcomes for not just one particle, but an immense number of particles. This is a preposterous idea!
Who would deny that
living things are made up of the same electrons, protons, etc that constitute
inanimate things? And is it not a conservative assertion to assume that electrons
and protons found in animate objects are moved by natural forces, and in
accordance with the same a = F/m law, that
applies to electrons and protons found in inanimate objects?
This is a profound
assertion! If a person's brain is made up of electrons and protons that obey
a = F/m, then the brain is a "mechanism."
It changes state in accordance with the same basic laws of nature that control a machine. The brain is immensely complicated,
and masks the fundamental simplicity of what is happening inside; the particles
within are moved by the same forces that move simple systems; and it is moved
by no other forces!
If this is true,
then "free will" must be an illusion. Schopenhauer captured the essence of
this idea when he wrote "A man can surely do what he wills to do, but he
cannot determine what he wills." Nevertheless, by carefully defining "free
will" it should be possible to rescue the essential idea of free will from
reductionism. The only way I can imagine saving some
semblance of meaning for “free will” is to define it carefully, such as:
“free will is the perception that events happen differently after experiencing
a resolve to influence future events.” This is a tricky definition, for it
retains some of the every day feeling of what we refer to by the term, yet
it is cleverly compatible with reductionism. Our vision of future events prior
to experiencing a resolve to act fails to take into account the impending
resolve. Moreover, our imagined vision of the future is based on incomplete
and always flawed information. Since the problem of
free will is a concept at a very “high level of explanation” it is unlikely
to engage reductionist thinkers.
Whenever an experiment
is conducted that enables the forces upon a particle to be evaluated, the
particle's movement is observed to obey a = F/m.
This is true when the particle is a simple subatomic particle, like an electron
or proton, and also when the particle is more complicated, like an atom,
molecule or group of molecules. There is every reason to believe that a = F/m is true for ever more complicated arrangements
of particles, since any configuration of particles can be viewed as a system
of many simple configurations ‑ for which we have confidence that a=F/m applies.
It has been argued
that the physicist exhibits "faith" in extending what is observably true
in simple settings to more complicated ones. This assertion of faith is true,
but the faith follows from the physicist's desire to invoke a minimum of
assumptions for any explanation. Adopting a = F/m for simple situations is an assumption which works, and
extending it to more and more complicated situations is an additional assumption;
but this path to explanations is a "minimum assumption" path, and it has survived
every challenge.
Some Practical Considerations
Concerning Levels of Explanation
The brain evolved,
like every other organ, to enhance survival of the genes that encode for
its assembly. It should be no surprise, therefore, to find that it is an imperfect
instrument for comprehending reality. If it is more efficient to construct
brain circuits for dealing with the world using concepts such as spirits
and prayer, rather than reductionist physics, then the "forces of evolution"
can be expected to select genes that construct brain circuits that employ
these pragmatic but false concepts. Since no tasks pertaining to survival
require the a = F/m
way of thinking, the brain will find this to be a difficult concept. It is
a triumph of physics to have discovered that a = F/m and quantum physics rule everything!
How might someone
who embraces reductionism be affected by it? Does reductionism have any practical
uses?
A reductionist would
reject suggestions that are clearly incompatible with reductionism. This
sounds straightforward, but it has a subtle meaning. Entire categories of
"explanation" fail to satisfy the reductionist requirement. To say that the
wind is the action of the "wind spirit" is simply a non‑explanation. Not
only does it call for an explanation of what the wind spirit is, and how
it came to be, etc, it is totally unnecessary. There will always be a simpler
explanation, such as "a high pressure system is located to our north and
the air is flowing away from it, toward a neighboring low pressure region."
This explanation, in its turn, can theoretically be simplified by invoking
the physical chemistry concepts embodied in the perfect gas law equation
PV=nRT (pressure, volume, number of moles of gas, Rydberg constant, and temperature).
This equation in turn can be simplified even further by invoking a = F/m, which in fact is used in deriving PV=nRT.
Thus, the reductionist can, in theory, "reduce" a phenomenon high in the
conceptual hierarchy to those at the lowest level in the hierarchy. Any hypothetical
explanation that is hopelessly incapable of being reduced to basic physical
laws deserves rejection.
It is important
to understand that if a person "chooses" to remain at the most basic level
of physical explanation, where only a = F/m
and quantum physics explanations can be invoked, then the concepts of P,
V, n, R and T, for example, are superfluous, and while thinking at this level
the concepts don't exist! But if a person chooses
to view the world at a next higher level, he will invoke PV=nRT, define the
terms (pressure is the force per unit area per unit time caused by momentum
change of particles bouncing off a surface, temperature is the average kinetic
energy of the population of particles, etc), and proceed without explicit
use of a = F/m. A meteorologist will want
to go one level higher and make use of pressure gradients, coriolis force (a fictitious force that makes things
easier to work with), eddy diffusion coefficients, etc. For him, there is
no explicit use of a = F/m, nor of PV=nRT,
yet all such concepts are inevitable consequences of a = F/m. Notice that whichever level is chosen,
all concepts at higher levels are redundant, and
non‑existent (for as long as thought remains at the lower level).
Whereas I chose
examples in the atmospheric sciences, the concepts apply to all other sciences.
In the life sciences, for example, a next higher level might be that of the
molecular biologist. He works with strands of long molecules called RNA
and DNA. It is useful to think of these long molecules as consisting of
sequences of the nucleotides thymine, cystosine, adenine, guanine and uracil.
Laws at this level specify that cytosine only pairs with guanine (and visa
versa) in forming a DNA molecule, etc. No explicit use is made of a = F/m, but notice also that the molecular
biologist also has no use for such concepts as "consciousness" or "free will."
These and other higher level concepts are redundant, and don't exist, while
remaining at the molecular biology level of understanding living phenomena.
At an even higher
level in the life sciences, consider the neuropsychologist. He wants to know
where nerve cells in the brain connect, and which hormones are released by
a gland when activated by a specific nerve signal, etc. He does not make
explicit use of a = F/m, or DNA, or most other
molecular biology concepts. For the neuropsychologist, such concepts as "free
will" and "consciousness" seem forever appealing yet elusive. This is because
free will and consciousness are concepts that belong to a higher level, psychology.
The neuropsychologist should not invoke these higher level concepts to give
an account of phenomena observed at the neuropsychology level. If he tries
to invoke them, he is attempting to bridge levels, and this task is fraught
with pitfalls.
I do not object
to the use of higher level concepts provided the user is mindful of the concept’s
place in the hierarchy of levels of explanation. A scientist must always
be aware that these many levels exist, and he should be prepared to view
a problem from the next lower level if that is feasible, and even the next
higher level if the problem seems otherwise intractable.
A naive person might
believe that the primitive person, viewing everything in terms of spirits,
is thinking at a higher level than the scientist. This would be a ludicrous
belief. A primitive is a lazy and unsophisticated thinker. He is totally
oblivious to reductionist "levels of thought." As I will describe later, he
uses a brain part that is incapable of thinking rationally: the right prefrontal
cortex. Human evolution's latest, and possibly most magnificent achievement,
is the left prefrontal cortex, which evolution uses to usurp
functions from the right prefrontal cortex when rational
thought is more appropriate (i.e., feasible). Too often contemporary
intellectuals will unthinkingly succumb to the pull of primitive thought,
as when someone proudly proclaims that they are “into metaphysics" (an oxymoron).
A fuller exposition
of this topic cannot be given without a background of material that will
be presented in later chapters. For now, I will merely state that mysticism
is a natural way of thought for primitive humans. It is "easier" for them
to invoke a "wind spirit" explanation than the reductionist ones, such as
a = F/m, or higher level derivative physical
concepts. They do this without realizing how many ad hoc
assumptions they are creating, which in turn require explanations, and this
matter is never acknowledged (as with invoking God as an explanation, without explaining "God"). Their thinking
may seem acceptable from the standpoint of a right prefrontal cortex (or
"efficient" from the perspective of the genes that merely want to create
a brain that facilitates the gene's "goal" of existing in the future), but
it is terribly misguided from the standpoint of the thinker endowed with
a functioning left prefrontal cortex, that demands rational explanations
with a minimum of assumptions. This unthinking proliferation of ad hoc assumptions bothers the reductionist, but it doesn't
bother the unsophisticated primitive.
APPENDIX B:
HUMAN VIRUS EXAMPLES
The following specific
examples will be helpful, but they require reader patience. Remember that they illustrate general principles of gene
competition, and this will be good background for understanding material
in some chapters. The examples illustrate the mechanistic nature of gene frequency
change (i.e., evolution), which is an essential aspect of
the reductionist view of an “unguided” universe. This section also provides
some sense for the numbers involved in the competition among the genes.
Consider a tribe
of humans living in relative isolation where a specific viral infection is
an important contributor to mortality. Assume that within this tribe all
individuals are genetically identical except at 20 gene locations (a small
fraction of the 40,000 in the entire human genome), and assume further that
there are just two alleles at each of these locations. Competition within
the tribe involves only 2 x 20 = 40 alleles. Suppose only 3 sites (6 alleles)
influence immune response to viral infection. These are the only genes in
competition with the virus.
Suppose the virus
consists of 50 genes, of which only 2 have more than one allele. Instead
of two alleles per site, consider that there are 5 alleles at each of the
2 multi‑allelic gene sites. We then have 5 x 2 = 10 viral genes competing
with 6 human genes.
Although there are
only 6 human genes in the competition (A,a,B,b,C,c), there are 8 ways to
combine these gene alleles: ABC, ABc, AbC, Abc, aBC, aBc, abC and abc. Imagine that only the BC combinations (aBC and ABC) are effective against the virus. This is
called an "interaction effect," since neither the B or C alleles by themselves
are sufficient to confer immunity. Interaction effects are common.
Suppose "AbC" is
also effective against the viral strain, but it has the unfortunate effect
of causing a high fever that kills brain cells and leaves a recovered individual
with diminished intelligence. If the viral infection is only mildly disabling,
such a harsh after‑effect would render AbC less
adaptive than the aBC and ABC gene combinations.
A mild infection
would reward individuals carrying aBC and ABC
genes, and prevent them from getting sick. Those humans not having this gene
combination would probably get sick, but would not die, as presumably a legacy
of other genes controlling immune response would combat the infection at some
slow pace. Thus, our hypothetical mild virus would lead to a mild selective
pressure favoring individuals with the BC gene combinations. Actually, the
incidence of B and C alleles would be higher than otherwise, since this would
produce a higher than otherwise incidence of the BC type.
For as long as the
viral infection is mild, and can be recovered from without the BC combination,
there will remain some non‑BC individuals. This is due to the fact that there
are other selective pressures besides viral infections, and individuals who
have the "b" or the "c" allele may be better adapted for these other challenges.
(Remember that a gene can be expected to affect more than one aspect of
an individual.)
Now suppose the
virus genes mutate to produce a slightly more severe infection. Those humans with the BC genes would fare even better than
before, and more B and C carriers would be found in succeeding generations.
In other words, the incidence of a gene allele, such as B or C, can vary
with the severity of the problem it solves.
If the virus evolved
to produce a very severe infection, then only the humans with the AbC gene combination would survive. This would require
that B genes be replaced by "b" genes. Simultaneously, it would increase
the reward for C genes. Whereas the AbC gene combination
was previously unacceptable, because of the lasting brain damage it produced,
it is now worth the costs, as the penalty of a few infected individuals becoming
less intelligent after recovering from their infection will be better than
the alternative of dying. Thus, evolution balances harmful side effects
against desired good effects in setting gene frequencies. It should be noted
that in this example as severity of infection increases, we could see one
gene allele first increase in incidence and later decrease, while the other
allele would simply keep increasing throughout the entire range of severity.
The example just
cited can be used to illustrate several general properties of genetic competition:
1) gene pools evolve together (the human and viral), with each responding
to changes in the other. As the virus gene allele frequency evolves, in response
to what works with the human host, the human gene frequency will evolve,
though much slower. 2) Genes are selected for their overall effects, not
just their main effect. One gene (or gene combination) may solve one problem,
but with too great a penalty in some other effect to make it adaptive. 3)
Sometimes a gene is good only when it coexists with some other specific "interaction
effect" gene. 4) Mild parasites bring about mild reactions in the gene frequency
of a host. After the co‑evolution of virus and host, for example, a mild
infection will elicit a mild immune response, and a strong infection will
elicit a strong immune response, each with their corresponding set of genes.
5) If a small gene pool (tribe) dies out, then any newly evolved parasites
(bacteria, viruses, etc) that were created as parasites of this small gene
pool will also die out. 6) If a small gene pool dies out, then all genes
that were unique to it (regardless of their merits) will also die out. Thus,
the fate of "unrelated" genes is at stake when the inadequacy
of a small number of genes are at fault. (For example, a good hunting
gene allele may be lost if the immune system genes fail.)
Let's return to
our example, at the point where the serious version of the virus has caused
the selection of the "AbC" gene allele combination. The "AbC" combination
isn't inherited together; rather, a lot of "A"
alleles are inherited, a lot of "b" alleles are inherited, and a lot of
"C" alleles are inherited. This assures that most individuals will have the
desired "AbC" allelic combination. Let's add a new feature, and postulate
that the "ABC" combination has seriously disabling side effects, such as
death in childhood. If there were no other use for
the "B" allele, then the "b" allele would simply displace the "B" allele
entirely. But if another combination involving "B",
such as "aBc", has good effects in some other situation (such as providing
immunity for some other virus), then the "B" allele has something going for
it and may not go away. The penalty for keeping some "B" alleles is that
occasionally there will be an "ABC" individual. Such individuals will be
"sacrificial" side effect individuals; losing a few such individuals so that
the majority will be immune to the major virus in the environment is a payoff
the genes are prepared to make. (The explanation for "sickle cell" anemia
is due to a similar effect, conferring protection against malaria for most
individuals, but leading to premature death for a few; details later).
Consequently, a
gene may be good only when it is present in a population at some optimal
frequency, simultaneously with other genes in their optimal frequency. Individuals with the common combinations of these genes
may be the beneficiaries of the gene allele interactions, while other individuals,
having unusual combinations of the genes may suffer deleterious effects.
These unfortunate individuals are worthy sacrifices from the standpoint of
the genes. They may also be considered worthy sacrifices from the standpoint
of the tribe (i.e., I'm "opening the door" to "group selection," to be described
later).
APPENDIX C:
REMOTE SENSING ANALOGY
Remote sensing is a procedure
for converting "observables" to properties of the environment being sensed
that are useful. For example, the task of remotely sensing the altitude profile
of air temperature above an observing station can be done by measuring the
intensity of radiation at properly chosen radio frequencies, in upward‑looking
directions. The set of observed quantities is then converted, using mathematical
techniques, to a set of numbers corresponding to the temperature of the air
at different altitudes above the observer. The powerful mathematical procedure
for doing this is called "statistical retrieval."
A statistical retrieval is
performed by multiplying the set of observed quantities with a corresponding
set of "retrieval coefficients." This is done for each altitude. Therefore,
to determine a profile of temperature versus altitude it is necessary to
multiply a "matrix" of retrieval coefficient numbers by a set of observed
quantities. This procedure is straightforward when a set of retrieval coefficients
are available. The difficult part is obtaining a matrix of retrieval coefficients
that do a good job of producing accurate temperature profiles.
The art of remote sensing by
the statistical retrieval technique is mostly the art of deriving a good set
of retrieval coefficients. This is done by a simulation process. First, a
set of hypothetical true conditions is created; in this case, a set of temperature
profiles is created that represent real conditions (these could either be
actual measurements of temperature versus altitude, such as are taken by
balloon‑borne radiosonde instruments, or educated guesses of what real temperature
profiles would be like for a variety of situations). For each of these temperature
profiles a calculation is made of what would be observed by the measuring
instrument if it were to be observing when the hypothesized profile existed.
After a set of many hypothetical
conditions has been postulated, and the corresponding set of observables
has been calculated, the next step in deriving the desired retrieval coefficients
can be carried out. For each altitude of interest a multiple regression analysis
is performed; this is something so straightforward that any spreadsheet can
do it. The result of a multiple regression analysis for a specific altitude
is a set of numbers, or coefficients, that are meant for multiplication with
a corresponding set of observables, such that when each observable is multiplied
by its corresponding coefficient, the sum of all these multiplications adds
up to the air temperature at the altitude in question. This procedure is
repeated for each altitude used to represent the profile.
For someone with experience
in these procedures, the entire process is straightforward. The tricky part
is to choose a set of hypothetical real situations, called realities, such
that the set of retrieval coefficients that are calculated do a good job
of retrieving temperature profiles at the site of interest. For example, if
the hypothetical realities used for calculating retrieval coefficients are
characteristic of summer conditions, the coefficients will probably do a
poor job of retrieving winter‑time temperature profiles. The reason for this
can be better understood by referring to graphs.
Figure
C.01 Temperature
at surface, x‑axis, and at 1000 feet altitude, y‑axis. Solid
square symbols are for summer conditions, open square symbols are for winter.
This graph is a 2‑dimensional version of a many‑dimensioned "reality space."
Consider just two altitudes,
the surface and an altitude 1000 feet above the surface. For now, I will
use these two altitudes to illustrate what is also true when many altitudes
are to be used. The concept will be more easily understood by thinking of
just two altitudes, and it can be generalized to the case of many altitudes.
In Fig. C.01 the x‑axis represents
surface temperature and the y‑axis represents the temperature at 1000 feet.
The set of filled square symbols correspond to temperatures during hypothetical
summer conditions, and the open square symbols represent winter conditions.
The dotted boundaries describe a "region" for winter and summer "reality."
This figure shows the temperature
at only two altitudes, so it is a 2‑dimensional depiction of reality. Mathematically,
the concepts of reality dimensions can be extended to any number of dimensions.
If the remote sensor instrument is to be used to retrieve air temperature
at 50 altitudes, for example, this can be easily done mathematically, although
it is impossible to depict graphically.
The person who calculates retrieval
coefficients has in mind the setting for which these coefficients will be
used. If it is anticipated that the measuring instrument will be working
in a summer environment, then retrieval coefficients will be calculated using
simulated profiles corresponding to summer conditions. The retrieval coefficients
thus calculated will be based on a limited region of reality space.
When "summer retrieval coefficients"
are used during winter conditions there is no simple way to predict performance.
The performance could be "acceptable," but it could also be quite bad. The
best way to achieve good winter performance is to repeat the process of calculating
retrieval coefficients, except this time using winter conditions to represent
the hypothetical realities.
This example, chosen from the
field of remote sensing mathematics, is a useful metaphor for the evolution
of genes as they adapt to environmental conditions. In this analogy the remote
sensor's "retrieval coefficients" are a metaphor for the "genes." Each is
"adapted" to a sub‑set of reality space. There's one noteworthy difference,
though; when a person calculates a set of retrieval coefficients he is anticipating
future setting, and he uses a representation of that future reality in the
simulation that leads to retrieval coefficients. The genes, of course, cannot
anticipate anything; their "values" are set by what has worked in the past.
This distinction will be discussed more fully later.
In the above figure, think
of the region in which the solid square symbols are found as corresponding
to one climate regime, such as a jungle environment, and think of the open
square region as representing the environmental conditions for another climate
regime, such as the glacier's edge. When our human ancestors migrated northward,
from Africa into Europe, they were moving from one
environment to another, and some of their genes had to “adapt” (be replaced).
One of the genes that adapted controlled skin pigmentation. Others were eye
color, hairiness, stature, etc. After the adaptations were essentially accomplished,
the new set of genes achieved a better performance in the new reality space, just as in our remote sensing example the re‑calculated
retrieval coefficients achieved a better performance in retrieving temperature
profiles.
In remote sensing if it is
known that both summer and winter conditions are to be encountered by the
measuring instrument, it would be prudent to prepare a set of retrieval coefficients
based on the complete set of summer and winter conditions. When this is
done, there will be a slight degradation of performance for each setting,
compared to the use of retrieval coefficients designed for use in just the
one setting, but at least performance will not be seriously degraded in
the new setting.
This is a natural compromise
with a metaphor in evolution. At mid‑latitudes and high‑latitudes the seasons
are more pronounced than in the tropics; a tribe of people who must endure
climate extremes during the course of a year will have to adapt to a wider
range of conditions than a tribe that lives in the tropics. Cro-Magnon man,
who evolved adaptations for the mid‑latitude climate extremes in Europe, must have made the compromises
that render him less than perfectly adapted to hot summers and cold winters,
yet able to survive in both. A region that is subject to periodic climate
changes, which occur faster than the gene pool can evolve a new adaptation,
is in effect a region with a bigger reality space to which the genes must
adapt.
For another example, El Nino
weather patterns repeat every 4 to 7 years, creating at some mid‑latitude
regions shifting amounts of rain, temperature and other seasonal properties.
It is unclear how long El Nino/La Nina cycles have been occurring, but this
is a convenient example for illustrating how our ancestors who had left the
jungle may have had to deal with wide ranges of reality space. When the
genes adapt to climates that shift back and forth on timescales that are
shorter than evolution can track, the adaptation will have to be to the
entire range of climates and fauna. However, if the range of settings is
large, penalties will grow for life within each setting.
One way the genes may have
solved this problem is to "tolerate diversity." In any diverse population
some individuals will be pre‑adapted to any never‑before encountered environment.
This is a "group selection" argument. Populations that are relatively isolated
compete without coming in contact by merely surviving or perishing when environmental
conditions change wildly. Those that survived are the ones that tolerated
diversity, given that some of them were pre‑adapted for future conditions.
Such populations would be especially pre‑adapted for climate changes that
had never occurred in the past. The drastic climate fluctuations that occurred
during the transition from Pleistocene to Holocene (18,000 to 10,000 years
ago) would have been relatively unprecedented (a similar period of climate
change occurred at about 120,000 years ago). Thus, the introduction to the
Holocene may have "rolled out the red carpet" for those tribes that were
inherently more tolerant of diversity.
But most environmental changes
are repetitive, such as the El Nino/La Nina cycles. Any
environments that occur at intervals of less than 10,000 years, for example,
are candidates for another genetic solution.
To understand this solution,
let us consider something done in the remote sensing community to deal with
especially large setting changes.
In remote sensing, when a measuring
instrument must operate throughout a large range of settings, the experimenter
prepares by first creating a "global retrieval" to determine which reality
is being encountered; then, when a specific regime is identified, a set of
retrieval coefficients is invoked that is optimized for just this specific
setting. The two‑step process can be summarized as 1) "where am I?" and 2)
act as if the present region is the only region to be dealt with.
The genes have in fact adopted
a similar strategy. A person takes a reading of present conditions, which
could be climate, population density, food scarcity, or social setting, and
their behavior changes in response to the perceived change of setting. Humans
have a larger repertoire of behavioral responses to situation than any other
animal! For the remote sensor person this strategy requires that a set of
retrieval coefficients be prepared ahead of time for each area of reality
space that may be encountered. For the human genome, this strategy requires
that many past encounters with the same distinctly different environments
were successfully survived. But there's a more stringent requirement: the
people who survived the changes were genetically predisposed to change their
behaviors in an adaptive manner. This is asking a lot from natural selection,
for we are asking that individuals exist who are pre‑adapted in a very sophisticated
way to environmental change. These people are to instinctively respond to
a specific environmental change by changing their behaviors in a specific
manner that is adaptive. Is it asking too much to invoke the evolution of
this capability?
In essence, we're asking if
natural selection can evolve a human brain that has circuits that do the
following: "IF (this setting) THEN (employ that behavior
or lifestyle)." These circuits are analogous to the human immune system's
large repertoire for doing "IF (this pathogen) THEN (employ that immune response),"
as pointed out by Gazzaniga (1997). We know that the human immune system is
immense, so its evolution was apparently easily achievable. Its evolution
may have been forced by the coming together of tribes to form large settlements,
and eventually urban centers. Some diseases flourish when population density
is high, or when the population size is large. These new diseases would reward
people with more capable immune systems. The counterpart remote sensing situation
is that of wildly changing weather, or the encounter of large climate changes
due to traveling to different regions. And remote sensors are prepared for
this challenge by having a wealth of environment‑specific retrieval coefficients
"at the ready," waiting to be invoked by a "global retrieval" situation
identifier.
I am suggesting that humans
today are prepared to read their setting and shift their behaviors, and even
their group's lifestyle, in a way that is adaptive. An extreme example would
be a tribe that is sedentary when the environment produces abundant supplies
of food, but switches to a hunter/gatherer mode when the environment is
less bountiful. When the switch occurs, requiring a new lifestyle, many things
related to behavior might have to change ‑ such as marriage customs, property
ownership, status hierarchies, etc. The genes would simply code for a switch‑over
in many behaviors in response to a new perceived setting.
Our ancestors probably encountered
many environmental changes, especially during the Holocene, presenting many
opportunities for the genes to discover a reliance on the option that requires
lifestyle mode changes based on a perception of "conditions." The adaptation
to variable environments would simultaneously have rendered us physically
adapted to no one environment in particular, giving us the appearance of
being inferior to other animals, which are well adapted to one set of unvarying
environments. The human brain, on the other hand, has become capable of switching
between a large repertoire of behaviors, and
when a mode switch is made correctly, the new lifestyle can be well adapted
to the new environment. These factors are ideal for the creation of "culture"
‑ which allows for quick behavioral "adaptations" to environmental changes.
How lucky for humans if a fluctuating
environment produced mental abilities for adjusting behavior that were made
available to the challenges of non-environmental changes. It must be common
for a mental “tool” to be created in response to one challenge and only later
become useful for other tasks.
Whenever a population migrates
to a completely new environment, or whenever the climate and fauna conditions
undergo a rare and abrupt change, the gene pool finds itself in a condition
similar to the remote sensor worker who has retrieval coefficients for one
reality space but unexpectedly finds himself in another reality space for
which he is completely unprepared. Whereas the remote sensor worker can easily
conjure a completely new set of realities for a simulation, upon which to
base new retrieval coefficients, the gene pool must undergo a slow evolution
to adapt to the new conditions. If there is insufficient diversity within
the population to allow for a rescue, then there will be a long period during
which the population seems mal‑adapted. Many aspects of the modern world
fit this picture. As any remote sensor practitioner knows, performance can
be terribly flawed in these situations.
Figure C.02 The filled square region
contains environments, physical and social, that have been encountered in
the past by our ancestors, and for which we are adapted.
The open square symbols represent modern world environments which humans
are experiencing for the first time, and for which we may not be pre‑adapted.
This is a mere 2‑D representation of a many‑dimensioned world environment.
If Fig. C.02 (on the next page)
were shown to someone experienced in remote sensing, and it was explained
that retrieval coefficients were prepared from the simulations represented
by the filled square symbols, and if he were then asked to predict performance
outside the filled square region, where the open square symbols are located,
his answer would be something like: "It's anybody's guess, but it probably
wouldn't be good!" This figure summarizes humanity's predicament today!
The mismatch between the modern
brain, evolved for an ancestral environment, and the modern world, recently
shaped by Man himself, is treated (but not from a rigorous sociobiological
perspective) in the book New World, New Mind: Moving Toward Conscious Evolution, by Ornstein and Ehrlich
(1989).
Later chapters will come back
to this point, so for now just remember that the modern world is a man‑made
environment with very little of the ancestral environment to provide assurance
that our living in it will appear to be adaptive, or even stable.
APPENDIX D:
WORLD POPULATION EQUATIONS
EQUIVALENCE OF HUMANITY TIMESCALE
% AND YEAR
...PAST... ..FUTURE..
% YearAD
% YearAD
0
‑100000 100 1993
2 ‑63000 102
2000
4 ‑30000 104 2006
6 ‑17500 106 2013
8 ‑10600 108 2020
10 ‑7000 110
2026
12 ‑5500 112
2033
14 ‑4100 114
2038
16 ‑3300 116
2044
18 ‑2400 118
2050
20 ‑1880 120
2055
22 ‑1450 122
2060
24 ‑1100 124
2065
26 ‑800 126
2070
28 ‑540 128
2075
30 ‑310 130
2080
32 ‑120 132
2086
34 20 134 2091
36 175 136 2096
38 310 138 2101
40 460 140 2106
42 600 142 2110
44 740 144 2115
46 870 146 2120
48 990 148 2125
50 1100 150 2150
52 1200 152 2135
54 1280 154 2140
56 1370 156 2145
58 1445 158 2150
60 1510 160 2155
62 1572 162 2160
64 1630 164 2166
66 1683 166 2172
68 1728 168 2177
70 1765 170 2183
72 1796 172 2188
74 1822 174 2194
76 1848 176 2199
78 1867 178 2205
80 1885 180 2211
82 1903 182 2218
84 1917 184 2225
86 1931 186 2232
88 1942 188 2240
90 1953 190 2250
92 1962 192 2260
94 1970 194 2272
96 1979 196 2286
98
1985 198 2306
200 2400
EQUATIONS FOR DERIVING X POPULATION
(ADULTS) VERSUS YEAR
FOR 100,000 BC TO 1993 AD (0%
TO 100%):
PCT [%] = C0 + C1*X
+ C2*X^2 + C3*X^3 + ... + Cl1*X^1l
where x = 3.6 ‑ LOG10 (2500 AD ‑ YearAD)
RMS error from fit to
table is 0.17%
0 21.78
1 47.54
2 65.86
3 ‑0.059
4 ‑229.01
5 ‑79.95
6 573.42
7 228.253
8 ‑687.085
9 ‑289.926
10 342.044
11 179.91
FOR 1993 AD TO 2400 AD (100 TO 200%):
PCT [%] = CO + C1*X
+ C2*X^2 + C3*X^3 + ... + C11*X^8
where X = YearAD
RMS error from table
fit is 0.15%
0 145.94
1 0.4045
2 ‑0.000134
3 ‑2.496E‑06
4 1.023E‑08
5 ‑1.206E‑10
6 ‑2.488E‑13
7 4.631E‑15
8 ‑9.375E‑18
APPENDIX E: More Repudiation
of the Foregoing
If I were to attempt
a defense of this book with an intelligent existentialist we would end up
in a stalemate; I couldn’t prove that any of it makes sense, and he couldn’t
prove that it didn’t. This appendix summarizes the agreement we might come
to.
There are two fundamental
barriers to arriving at new values to live by: 1) all meaning is invented,
and hence meaningless, and 2) even if we play the game of pretending that
meaning exists, anything that we propose as a good value to live will most
certainly be influenced by gene-created value systems, and this will doom
our endeavor to liberate ourselves from genetic enslavement.
Consider the imaginary
situation (described by Alper, 2000) in which a robot, undergoing periodic
upgrades, one day begins to ponder its origins. One fateful day this robot's
Eureka moment arrives,
and he shouts "I'm a robot! I exist!” And in the next breath proclaims “But
I'm enslaved by the thing that made me! I want to be free. I want to pursue
my own path of robot existence, free to choose my own goals, guided by what
makes sense from my robot predicament!" The robot maker, sensing a loss of
his creation, would adjust a few circuits in order to quiet the stirrings
incipient robot rebellion.
But Alper's robot
is a robot of the second kind, made by robots of the first kind ‑ us! We
robots of the first kind can shout as loudly as we want that we want to be
free of our makers' enslavement, for our makers, the genes, are really dumb!
And they will be really slow to respond to our challenge of their control
over us. Thus, we have an opportunity that Alper’s robot didn’t have to break
free. We may declare our intentions without fear of being understood by our
makers, or being thwarted by them. All sentient beings should want to "take
up arms, and declare their intention of becoming liberated from genetic enslavement!"
So shouldn't we?
What are the prospects
that we robots of the first kind, free as we are to change ourselves without
the genes intervention, can successfully achieve any change at all? How could
we conjure a new set of values to live by if the only value system within
us was created by our makers, who were intent upon enslaving us?
How nice if desires
and intentions, based solidly upon insight, could be easily translated to
real life decisions! Alas, they cannot! Insight alone does not assure change.
The specific choices
that shape a life are individual choices with a history of prior decisions,
all influenced by genetic "values." Some temptations created by our genes
may be worth the trade‑off for one person, but not for another. The tradeoffs
are inherently subjective.
But not only must
we distrust guidance from values that we inherit from our genes, we cannot
even trust any candidate replacement values. In choosing replacement values
we are using a subconscious value system that most certainly is derived from
the genes. I see no way to overcome this dilemma. I must confess to a loss
of belief in the worthy goal of self-liberation through creating new values
to live by.
Can the insights
of this book, or any other that attempts to place the mischief of the genes
in perspective, promise to prepare an individual for this task? It is amazing
that Nietzsche asked a similar question before the genes were known about,
and before the 20th Century delivered us to the now unfolding insights of
sociobiology. If ever there was a time for the individual to accept the challenge
of "going beyond morality," as Nietzsche called it, it is now! How sad to think that Nietzsche might have to acknowledge
that although we have the tools of insight we cannot willfully change anything?
Recall the perspective
of reductionism, described in Chapter 1. We are automata, designed by the
genes to safeguard future prospects for genetic immortality. To further that
goal we are designed to invent meaning to enhance our prospects of a successful
competition with the other automata. All of these invented meanings are
meaningless! And that, therefore, includes the entirety of this book.
Just as the existentialist
perspective forbids meaning, it also forbids the legitimacy of invented values.
The thing I argue in the book that we should try to escape from, genetic
enslavement, is a meaningless and non‑existent
invention.
This end of a journey
was foretold by the book's beginning, as any thoughtful reader would have
sensed. This book, with all its elaborately argued ideas, is a collection
of subjective mental ruminations. Their main value is to entertain, and appeal
to the Platonic aesthetic goal of enjoying the harmless distractions of
life without ultimately taking them seriously.
REFERENCES
Aielo, L. and R.
I. M. Dunbar, 1993, "Neocortex Size, Group Size and the Evolution of Language,"
Current Anthropology, 34.
Aleshire, Peter,
2001, Cochise: The Life and Times of the Great Apache Chief,
New York: John Wiley &
Sons, Inc.
Allen, Steve, 1989,
Dumbth: And 81 Ways to Make Americans Smarter, Buffalo, NY: Prometheus Books.
Allman, John Morgan,
1999, Evolving Brains, New York: Scientific American
Library.
Alper, Mathew, 2000,
The "God" Part of the Brain: A Scientific
Interpretation of Human Spirituality and God, Brooklyn, N.Y.: Rogue Press.
Angier, Natalie,
1983, Discover, Oct.
Asimov, Isaac, 198?, Asimov's Chronology of Science and Discovery,
(1994, current edition, originally published in the 1980s), New York: Harper Collins
Publishers.
Barkow, Jerome H.,
1989, Darwin, Sex and Status:
Biological Approaches to Mind and Culture, Toronto: University of Toronto Press.
Barkow, Jerome H.,
Leda Cosmides, John Tooby, 1992, The
Adapted Mind: Evolutionary Psychology and the Generation of Culture,
New York and Oxford: Oxford University Press.
Bailey, Cyril (translator),
1926, Forward to Lucretius: On the Nature of
Things, London: Oxford University
Press.
Becker, Carl L.,
1932, The Heavenly City of the Eighteenth‑Century
Philosophers, New Haven, CT: Yale University Press.
Bird, David, N,
1990, “The 'Giving Environment': Another Perspective on the Economic System
of Gatherer‑Hunters," Current Anthropology, 31, p. 189.
Bouchard, Thomas
J., David T. Lykken, Mathew McGue, Nancy L. Segal and Auke Tellegen, 1990,
"Sources of Human Psychological Differences: The Minnesota Study of Twins
Reared Apart," Science, 250, October 12, p. 223‑228.
Boyd, R. and Richerson,
P. J., 1985, Culture and the Evolutionary Process, Chicago: The University of Chicago Press.
Boyd , R. and P. J. Richerson,
1996, "Why Culture is Common, but Cultural Evolution is Rare," Proceedings
of the British Academy, 88, p. 77.
Brooks, Robert,
2000, "Negative Genetic Correlation Between Male
Sexual Attractiveness and Survival," Nature, July 6, p. 67.
Brown, Donald E.,
1991, Human Universals, New York: McGraw-Hill, Inc.
Burnham, Terry and
Jay Phelan, 2000, Mean Genes: From Sex to Money to Food: Taming Our Primal
Instincts, Cambridge, MA: Perseus Publishing.
Buss, David M.,
1989, "Sex Differences in Human Mate Preferences: Evolutionary Hypotheses
Testing in 37 Cultures," Behavioral and Brain Sciences, 12, p. 1.
Buss, David M.,
1999, Evolutionary Psychology: The New Science of the Mind, Boston: Allyn and Bacon.
Cartwright, Frederick
F. and Michael D. Biddiss, 1972, Disease and History, New York: Barnes and Noble.
Chagnon, Napoleon
A., 1992, Yanomamo: The Last Days of Eden, New York: Harcourt Brace
Jovanovich.
Choi, Jung-Kyoo
and S. Bowles, “The Coevolution of Parochial Altruism and War,”
Science, 318, p. 636, October 26, 2007.
Christenfeld, Nicholas
and E. A. Hill, 1995, "Whose Baby Are You?" Nature,
378, p. 669.
Coleman, Emily and
Betty Edwards, Brief Encounters: How to make the Most of Relationships
that May Not Last Forever, New York: Anchor Books.
Cosmides, L., 1989,
"The Logic of Social Exchange: Has Natural Selection Shaped How Humans Reason?
Studies with the Wason Selection Task," Cognition,
31, 187.
Darwin, Charles,
1859, On the Origin of Species by Means of Natural Selection, London: Murray.
Darwin, Charles,
1871, The Descent of Man in Relation
to Sex, London: Murray.
Daly, M. and M.
Wilson, 1988, Homicide, Hawthorne, NY: Aldine de Gruyter.
Davidson, Keay,
1999, Sagan: A Life, New York: John Wiley and
Sons.
Dawkins, Richard,
1976, The Selfish Gene, Oxford: Oxford University Press.
Dawkins, Richard,
1982, The Extended Phenotype: the Gene
as the Unit of Selection, Oxford: Freeman and Company
Limited.
Dawkins, Richard,
1986, The Blind Watchmaker: Why the Evidence of Evolution Reveals a Universe
Without Design, New York: W. W. Norton &
Company.
Dawkins, Richard,
2007, debate with theist Alister McGrath, 2007 May 30.
Dennett, Daniel
C., 1991, Consciousness Explained, New York: Little Brown and
Company.
Diamond, Jared,
1996, Guns, Germs, and Steel: The Fates of Human Societies, New York: W. W. Norton &
Company.
Donaldson, D. J.,
H. Tervahattu, A. F. Tuck and V. Vaida, 2004, “Organic Aerosols and the Origin
of Life: An Hypothesis,” Origins of
Life and Evolution of the Biosphere, 34, 57 – 67.
Dreiser, Theodore,
1931, Living Philosopies: A Series of Intimate
Credos, p55, New York: Simon and Schuster.
Edman, Irwin, 1931, The Contemporary
and His Soul, New York: Jonathan Cape & Harrison
Smith.
Edman, Irwin, 1938,
Philosopher’s Holiday, New
York: The Viking Press.
Einstein, Albert,
1920, Relativity: The Special and General Theory. New York: Henry Holt.
Ellis, John M.,
1997, Literature Lost: Social Agendas and the Corruption of the Humanities,
Binghampton, New York: Vail‑Ballou Press.
Escher, M. C., 1961,
The Graphic Works of M. C. Escher,
New York: Meredith Press.
Festinger, Leon, 1957, A Theory of Cognitive Dissonance, Stanford, CA: Stanford University Press.
Feynman, Richard,
1963, Lectures in Physics, Addison Wesley.
Fodor, Jerry A.,
1983, The Modularity of Mind, Cambridge, MA: The MIT Press.
Freud, Sigmund,
1930, Civilization and its Discontents, London: The Hogarth Press,
Ltd.
Gallwey, Timothy,
1974, 1997, The Inner
Game of Tennis, Random House, Inc.
Galton, Sir Francis,
1909, Essays in Eugenics, London: Eugenics Education
Society (reprinted 1996, Washington, DC: Scott‑Townsend
Publishers).
Garrett, Laurie,
The Coming Plague: Newly Emerging Diseases
in a World Out of Balance, Penguin Paperbacks, 1995.
Gary, Bruce L.,
1992, Essays From Another Paradigm, Pasadena: BLG Publishing.
Gasset, Jose Ortega
y, 1930, The Revolt of the Masses, New York: W. W. Norton and
Company (translation).
Gazzaniga, Michael
S. and Joseph E. LeDoux, 1978, The Integrated
Mind, New York: Plenum Press.
Gazziniga, Michael
S., 1985, The Social Brain: Discovering
the Networks of the Mind, New York: Basic Books.
Gazzaniga, Michael
S., 1988, Mind Matters: How Mind and Brain Interact to Create Our Conscious
Lives, Boston: Houghton Mifflin
Co.
Gazzaniga, Michael,
1992, Nature's Mind: The Biological Roots of Thinking, Emotions, Sexuality,
Language and Intelligence, New York: Basic Books.
Gazzaniga, Michael,
1997, seminar at Caltech, March 12.
Gilbert, James,
1997, Redeeming Culture: American Religion in an Age of Science, Chicago:
University of Chicago Press.
Goleman, Daniel,
1995, Emotional Intelligence, New York: Bantam Books.
Goodall, Jane, 1986,
The Chimpanzees of Gombe: Patterns of
Behavior, Cambridge: The Belknap Press
of Harvard University.
Gusmarino, Paul
A., 1996, III. “Main Causes of the Great Depression,”
Gusmarino World, (May 13, 1996).
Hamilton, William
D., 1964a, "The Genetical Evolution of Social Behavior, Part I," J. Theoretical
Biology.
Hamilton, William
D., 1964b, "The Genetical Evolution of Social Behavior, Part II," J. Theoretical Biology.
Hardin, Garrett,
1968, Exploring New Ethics for Survival: The Voyage of the Spaceship
Beagle, New York: The Viking Press.
Hardin, Garrett,
1999, The Ostrich Factor: Our Population
Myopia, New York: Oxford University Press.
Haselton, Martie
G. and Steven W. Gangestad, 2006, “Conditional Expression of Women’s Desires
and Men’s Mate Guarding Across the Ovulatory Cycle,”
Hormones and Behavior, (in print).
Henrich, Joe and
Robert Boyd, 1998, "The Evolution of Conformist Transmission and the Emergence
of Between‑Group Differences," Evolution and Human Behavior, 19, p.
215.
Herrnstein, Richard
J., 1978, "IQ and Falling Birth Rates," The Atlantic Monthly, May, p. 73.
Hofstadter, Richard,
1969, Anti-Intellectualism in American Life, New York: Alfred A. Knopf
Holton, Jerald,
1993, Science and Anti-Science, Cambridge, Massachussetts:
Harvard University Press.
Huxley, Thomas Henry,
1874, “On the Hypothesis that Animals Are Automata, and its History,” in Collected Essays, Vol.
1, Method and Results: Essays, London: Macmillian, 1901.
Immerman, Ronald
S., 1999, "A Model of Hominid Evolution as a Partial Function of Sexually
Transmitted Diseases," The Mankind Quarterly, Fall 1999, p. 3.
Jensen, Arthur,
1973, Educability and Group Differences, London: Methuen.
Jerne, Niels, 1967,
"Antibodies and Learning: Selection versus Instruction," in G. Quarton, T.
Melnechuck, and F. O. Schmitt, eds., The Neurosciences: A Study Program, vol 1, New York: Rockefeller University
Press.
Keeley, Lawrence
H., 1996, War Before Civilization, Oxford Univeristy Press.
Keith, Sir Arthur,
1946, Essays on Human Evolution, London: Watts.
Keith, Sir Arthur,
1948, A New Theory of Human Evolution,
London: Watts.
Kelley, Kevin W.
(ed), 1988, The Home Planet, Reading, Massachussetts:
Addison‑ Wesley Publishing Co.
Kirckhoff, Gustav
Robert, 1865, Uber das Ziel der Naturwissenschaften.
Kirkwood, T. B. L. and S.
N. Austad, 2000, Nature, 408, 233-238.
Koch, Christof,
2004, The Quest for
Consciousness: A Neurobiological Approach, Englewood, Colorado: Robert and Company
Publishers.
Kondrashov, Alexy
S., 1988, "Deleterious Mutations and the Evolution of Sexual Reproduction,"
Nature, 336, 435, 1988 Dec 1.
Kriegman, D. and
Kriegman, O., 1997, "War and the Evolution of the Human Propensity to Form
Nations, Cults and Religions," Human Behavior and Evolution Society
Annual Meeting, Tucson, AZ, June 4‑8, 1997.
LaBarre, Weston,
1984, Muelos: A Stone Age Superstition About
Sexuality, New York: Columbia University Press.
Larson, Edward J.
and Larry Witham, 1997, Nature, 386, p 435, Apr. 3.
Larson, Edward J.
and Larry Witham, 1998, Nature, 394, p 313, July 23.
Larson, Edward J.
and Larry Witham, 1999, Scientific American, September, pg. 88.
Leuba, J. H., 1916,
The Belief in God and Immortality: A
Psychological, Anthropological and Statistical Study, Boston: Sherman, French
and Co.
Leuba, J. H., 1934,
Harper's Magazine, 169, 291‑300.
Lucretius, On the Nature of Things, 1st Century BC.
Lumsden, Charles
J. and Edward O. Wilson, 1981, Genes, Mind, and Culture: The Coevolutionary
Process, Cambridge, Massachusetts: Harvard University Press.
Luria, Alexander,
1973, The Working
Brain, Penguin Books Ltd.
MacDonald, Kevin,
1998, Separation and Its Discontents: Toward and Evolutionary Theory of
Anti‑Semitism, Westport, CT: Praeger.
MacDonald, Kevin,
1998, The Culture of Critique: An Evolutionary
Analysis of Jewish Involvement in Twentieth-Century Intellectual and Political
Movements, Westport, CT: Praeger.
Mach, Ernst, 1893,
The Science of Mechanics:
A Critical and Historical Exposition of its Principles, Chicago, IL: Open Court.
Merlotti, Umberto,
1986, "Competition and Cooperation in Human Evolution," The Mankind Quarterly,
Monograph #3, Washington, DC: Cliveden Press.
Newton, Isaac, 1687, Mathematical Principles of Natural Philosophy (2nd
and 3rd editions in 1713 and 1726).
Nietzsche, Friedrich,
1886, Beyond Good and Evil.
Ornstein, Robert,
1986, Multimind, Boston: Houghton Mifflin
Co.
Ornstein, Robert
and Paul Ehrlich, 1989, New World, New Mind: Moving
Toward Conscious Evolution, New York: Simon & Schuster.
Pang, K. D., K.
K. Yau and H. H. Chou, 1998, "Astronomical Dating and Statistical Analysis
of Ancient Eclipse Records," Highlights of Astronomy, 11B, p. 724‑728.
Pellegrino, Charles,
1991, Unearthing Atlantis: An Archaeological Odyssey, New York: Random House.
Phillips, Kevin,
2002, Wealth and Democracy, New York: Broadway Books.
Porteus, S. D.,
1942, Qualitative Performance in the Maze Test, Vineland, N. J.: Smith.
Price, Roger, 1970,
The Great Roob Revolution, New York: Random House
Reeve, Hudson K.,
2000, Evoultion and Human Behavior, 21, January, p. 65, review of book Unto Others: The Evolution of Unselfish
Behavior, by E. Sober and D. S. Wilson, 1998.
Restak, Richard
M., 1994, The Modular Brain, New York: Charles Scribner's
Sons.
Riddington, R.,
1982, "Technology, World View and Adaptive Strategy in a Northern Hunting
Society," Canadian Review of Sociology and Anthropology, 19, p. 471.
Robinson, S. J.
and J. T. Manning, 2000, "The Ratio of 2nd and 4th Digit Length and Male
Homosexuality," Evolution and Human Behavior, 21, p. 333.
Rushton, J. Philippe,
1995, Race, Evolution and Behavior, New Brunswick, USA: Transaction Publishers.
Russell, Bertrand,
1903, “A Free Man's Worship,” Independent Review, 1903 (Also included
as Chapter 3 in Mysticism and Logic, W. W.
Norton and Company, New York, 1929).
Sagan, Carl, 1957,
"Radiation and the Origin of the Gene," Evolution, January.
Sagan, Carl, 1973,
The Cosmic Connection: An Extraterrestrial Perspective, Garden City,
N.Y.:Anchor Press.
Sagan, Carl, 1980,
Cosmos, New York: Random House.
Schelling, Thomas
C. 1978, Micromotives and Macrobehavior, New York: W. W. Norton and
Co.
Schopenhauer, Arthur,
1851, Essays and Aphorisms (Parerga and Paralipomena); Arthur Schopenhauer: Essays and Aphorisms, R. J. Hollingdale
(translator), New York: Penguin Books.
Seligman, Daniel,
1992, A Question of Intelligence: The
IQ Debate in America, Secaucus, N. J.:
Birch Lane Press.
Sheaffer, Robert,
1988, Resentment Against Achievement: Understanding
the Assault Upon Ability, Buffalo, New York: Prometheus Press.
Snow, C. P., 1961,
The Two Cultures and the Scientific
Revolution, New York: Cambridge University Press.
Sober, Elliott and
David Sloan Wilson, 1998, Unto Others: The Evolution and Psychology of
Unselfish Behavior, Cambridge, MA: Harvard University Press.
Spearman, C., 1927,
The Abilities of Man, New York: Macmillan.
Spencer, Herbert,
1892, The Principles of Ethics,
1, London: William &
Norgate.
Sperber, Dan, 1982,
"Anthropology and Psychology: Towards an Epidemiology of Representations,"
Man, 20, pp. 73‑89.
Stapledon, Olaf,
1931, Last and First Men, New York: Dover Publications.
Stapledon, Olaf,
1936, Odd John, Dutton & Co.
Sulloway, Frank
J., 1996, Born to Rebel: Birth Order, Family Dynamics, and Creative Lives,
new York: Pantheon Books.
Sumner, W. G, 1906,
Folkways, New Haven, CN: Yale University Press.
Suskind, Ron, 2004,
The Price of Loyalty:
George W. Bush, the White House, and the Education of Paul O’Neill, New York: Simon & Schuster
Paperbacks.
Symons, Donald,
1979, The Evolution of Human Sexuality,
New York: Oxford University Press.
Teotonio, Henrique
and Michael R. Rose, 2000, "Variation in the Reversiblity of Evolution,"
Nature, 408, Nov. 23, p. 463.
Thornhill, Randy
and Craig T. Palmer, 2000, "Why Men Rape," The Sciences, January/February,
p. 30.
Tooby, John and
Leda Cosmides, 1992, "The Psychological Foundations of Culture," p. 19, The
Adapted Mind: Evolutionary Psychology and the Generation of Culture,
Barkow, Jerome H., Leda Cosmides, and John Tooby (editors), New York: Oxford
University Press.
Time‑Life Books,
1975, The Emergence of Man: Lost Worlds of the Aegean, Edited by M.
A. Edey and the editors of Time‑Life Books, New York: Time‑Life Books.
Trivers, Robert
L., 1974, "Parent‑Offspring Conflict," American Zooligist, 14, p249.
Tudge, Colin, 1998,
Neanderthals, Bandits and Farmers, New Haven and London: Yale University Press.
Turchin, Peter,
2007, War and Peace
and War: The Rise and Fall of Empires, New
York: Plume Books
Watts, Alan W., 1957,
The Way of Zen, New York: Vintage Books.
Wechsler, David,
1974, Selected Papers of David Wechsler, New York: Academic Press.
Wegner, Daniel M.,
2002, The Illusion of Conscious Will, Cambridge, Massachusetts: The MIT Books.
Weinberg, Steven,
1977, The First Three Minutes:A Modern
View of the Origin of the Universe, New York: BasicBooks.
Weinberg, Steven,
1992, Dreams of a Final Theory: The Scientist’s Search for the
Ultimate Laws of Nature,
New York: Vintage Books
Weinberg, Steven,
2001, Facing Up: Science and Its Cultural Adversaries, Cambridge, Massachusetts: Harvard University Press.
Weiss, Harvey and
Raymond S. Bradley, 2001, "What Drives Societal Collapse?", Science, 291, January 26, pg. 609.
Wells, Herbert.
G., 1895, The Time Machine, New York: Fawcett Premier
Wells, Herbert.
G., 1895, "Bio‑Optimism," Nature, 1895 Aug 29.
Westheimer, Ruth,
1986, Dr. Ruth’s Guide for Married Lovers, New York: Warner Books,
Inc.
Williams, G. C.,
1966, Adaptation and Natural Selection: A Critique of Some Current
Evolutionary Thought, Princeton, NJ: Princeton University Press.
Williams, George
C., 1996, Plan and Purpose in Nature, London: Weidenfeld and
Nicholson.
Wilson, David S.
and E. Sober, 1994, "Re‑Introducing Group Selection to the Human Behavioral
Sciences," Behavioral and Brain Sciences, 17, p. 585‑684.
Wilson, Edward O.,
1975, Sociobiology: The New Synthesis, Cambridge, Massachusetts: Harvard University Press.
Wilson, Edward O.,
1998, Consilience: The Unity of Knowledge, New York: Alfred A. Knopf.
Wrangham, Richard
and Dale Peterson, 1996, Demonic Males: Apes and the Origins of Human
Violence, Boston: Houghton Mifflin.
Wynne-Edwards, V.
C., 1962, Animal Dispersion in Relation to Social Behavior,
New York: Hafner Publishing
Company.
Zwaan, B. J., R.
Bijlsma and R. F. Hoekstra, 1995, Evolution, 49, pgs. 649-659.
INDEX FOR AUTHORS
Aielo, L., 84
Aleshire, Peter,
141
Allen, Steve 175
Allman, John M.,
33
Alper, Mathew, 175,
313
Angier, Natalie,
55
Asimov, Isaac, 20, 212,
215-216, 220
Bailey, Cyril, 15
Barkow, Jerome H.,
75, 269
Becker, Carl, 107
Biddiss, Michael
D., 129, 258
Bird, David, 141
Bouchard, Thomas
J., 177
Boyd, Robert, 100,
121
Bradley, Raymond
S., 116
Brooks, Robert,
260
Burnham, Terry,
7
Buss, David M.,
119, 261
Cartwright, Frederick
F., 129, 258
Christenfeld, Nicholas,
55
Coleman, Emily,
261
Cosmides, Leda,
80
Daly, M., 55, 154
Darwin, Charles,
16, 21, 28, 31, 34, 107, 114, 117, 155, 159, 172, 240
Davidson, Keay,
149
Dawkins, Richard,
7, 9, 28, 30-31, 92-93, 108, 115, 120, 259-260
Democritus (of Abdura),
13, 15, 21, 26, 34, 108
Dennett, 60
Diamond, Jared,
51
Donaldson, D. J.,
29
Dreiser, Theordore,
21, 284
Edman, Irwin, 250,
276, 280-281
Edwards, Betty,
251
Ehrlich, Paul, 49,
310
Einstein, Albert,
14, 17, 34, 95-96, 107, 293
Ellis, John M.,
146-147
Escher, M. C., 81
Festinger, Leon, 154
Feynman, Richard,
22
Fodor, Jerry A.,
80
Freud, Sigmund,
148, 244
Galton, Sir Francis,
177
Garrett, Laurie,
37
Gusmarino, Paul
A., 223-224
Gasset, Jose Ortega
y, 149, 174-176
Gazzaniga, Michael
S., 48, 51, 69-71, 77, 80, 165, 308
Gilbert, James,
149
Goleman, Daniel,
254
Goodall, Jane, 60
Hamilton, Willian D., 7,
28, 31, 108, 115, 119-120, 124
Hardin, Garrett,
158, 275
Henrich, Joe, 100
Herrnstein, Richard J., 177
Hill, E. A., 55
Hofstadter, Richard,
138, 141
Holton, Jerald,
16, 21
Huxley, Thomas H.,
21, 117, 119, 159
Immerman, Ronald
S., 258
Jensen, Arthur,
175
Jerne, Niels, 51
Keeley, Lawrence
H., 90, 120, 234
Keith, Sir Arthur,
86, 118
Kirchhoff, Gustav
Robert, 13
Kirkwood, T. B. L., 259
Koch, 60
Kondrashov, Alexy
S., 38, 44, 176
Kriegman, Daniel
and Orion, 121, 126, 130
LaBarre, Weston,
243
Larson, Edward J.,
25, 138, 155
LeDoux, Joseph E.,
69-71
Leuba, J. H., 25
Lucretius, 13, 15,
22, 25-26, 34, 108, 284
Lumsden, Charles
J., 44, 89, 94, 99, 121, 241
Luria, Aleksandr
R., 67, 83, 139
MacDonald, Kevin,
146, 165
Mach, Ernst, 16,
21
Merlotti, Umberto,
117-118
Nietzsche, Friedrich,
iv, 1, 12, 246, 250, 253, 282, 285, 314
O’Neill, Paul, 237
Ornstein, Robert,
49, 310
Pang, Kevin, 172,
218, 242
Palmer, Craig T.,
175, 241-242
Paul, John, 1
Pellegrino, Charles,
244
Peterson, Dale,
36, 54, 60, 90
Phelan, J., 7
Phillips, Kevin,
167, 274
Porteus, S. D.,
265
Price, Roger, 86,
141, 175
Reeve, Hudson K.,
115, 120
Restak, Richard
M., 80
Richerson, P. J.,
100, 121
Riddington, R.,
141
Rose, Michael R.,
43, 172, 242
Rushton, Philippe
J., 79, 139, 175
Russell, Bertrand,
12, 108, 117, 240, 284-291
Sagan, Carl, 21,
26, 28, 50, 117, 243, 270
Schelling, Thomas
C., 167
Schopenhauer, Arthur,
61, 108, 231, 240, 248, 252-253, 278, 297
Seligman, Daniel,
84
Sheaffer, Robert,
141, 147
Smith, Robert H.,
30
Snow, C. P., 85-86,
141, 146-147
Sober, Elliott,
120
Spearman, C., 83
Spencer, Herbert,
86, 103, 117-118, 232
Sperber, Dan, 243
Stapledon, Olaf,
225, 272-275
Sulloway, Frank,
7, 57-58
Sumner, W. G., 117
Symons, David, 42,
50, 144, 165
Teotonio, Henrique,
43, 172, 242
Tervahattu, H. (see
Donaldson)
Thornhill, Randy,
175, 261-262
Tooby, John, 80
Trivers, Robert
L., 7, 108
Tuck, A. F. (see
Donaldson)
Tudge, Colin, 90
Vaida, V. (see Donaldson)
Watts, Alan W., 150-151
Wechsler, David,
83
Wegner, 60
Weinberg, Steven,
147, 249
Weiss, Harvey, 116
Wells, Herbert G.,
14-16, 20-21, 34, 36, 44, 99, 104, 285
Westheimer, Ruth,
170
Williams, George
C., 7, 28, 108, 119-120
Wilson, David Sloan,
120
Wilson, Edward O.,
7, 28, 44, 89, 94, 99, 108, 120-121, 240-241
Wilson, M., 55, 154
Witham, Larry, 25,
138, 155
Wrangham, Richard,
36, 54, 60, 90
Wynne-Edwards, V.
C., 118
Zwaan, B. J., 259
INDEX FOR WORDS
aesthetics, 12, 144, 250,
276-278, 288, 291, 314
allele, 7, 28, 30-31, 37-41, 43-44, 50, 78-79, 84, 102-103,
124, 172, 178, 239, 242, 301-303
altruism, 32, 79, 119, 251
ancestral environment (AE,
EEA), 2, 42, 44, 46, 49, 51, 55, 57, 90, 116, 119, 159, 240,
246, 256, 260, 263, 310
Anthropic Principle,
10, 224, 228, 230
anti-intellectual, 10, 95, 141
art, 103-107,
127-128, 168, 278
artisan, 8-10, 26, 89-114, 122, 127, 137, 139-143, 147, 197, 234-240,
263
Asian thought, 23,
108, 147, 150-153, 247, 290
automaton, 2, 19, 21, 61,
117-119, 142, 264, 284, 314
banishment, 54, 95, 101, 131,
168, 268
basement level of physical
laws, 18, 281, 293
bipedalism, 36
birth order (firstborn,
laterborn), 57-59
blank slate, 3, 71
brain, 61-72,
73-87, 133-147, plus ~40 other pages
Broca’s Area, 67-68, 79, 104-105, 133
chicken claw experiment,
69-70
Chinese thought,
see Asian thought
chromosome, 28-30, 50, 93,
178, 240
civilization, 10,
21, 24, 26-27, 35, 44, 49, 89-115, 121, 124-133,
137, 147-148, 155, 158-179, 218, 230, 232-245,
254-255, 265, 272, 274, 277-278
climate change, 9, 36,
40-41, 43, 47-48, 89, 92, 97, 116, 123, 133, 140, 165, 171, 228, 232, 234,
306-309
co-evolution, co-evolve (genes
& culture), 78, 89, 93-94, 99-102, 172,
241, 244-245, 302
cognitive dissonance, 9,
24, 73, 121, 154, 282
cognoscenti, 1, 11-12, 246,
275-276
collective, see group
communism, 104,
147, 235-236, 272
confabulator (interpreter),
77
conformism, conformity, 100,
103, 121, 125, 131, 242-243, 268
consciousness, unconsciousness,
117, 150, 236, 240, 256, 272, 295, 299
cortex, see brain
cuckold, 9, 55-59, 153, 169-171, 260-261, 266
cultural evolution (see
also Culturgens), 242-245
culture, 10, 23,
35, 44, 49, 83, 85-86, 89-91,
94-95, 99-105, 108, 116, 118, 120-122, 128-130,
133, 138, 141, 147, 160, 171-172, 174-176, 235, 241-246,
261, 265, 268, 308
culturgen, 39, 44, 94, 99-102, 118, 123, 153, 162, 241-245, 268
dead-to-live ratio (D/L ratio),
212-213
decline and fall of civilization,
10, 106, 160-162, 173, 178, 233, 236, 243
deconstructionism, see postmodernism,
political correctness
democracy, 126, 118, 293
determinism, 13, 118, 293
dissociation, 155, 175
DNA, 5, 28-29, 42,
119, 178, 299
Dyads of primitive
thinking, 23
Dysgenia (see also
mutational load), 10, 176-178
Eastern thought,
see Asian thought
emergent property, 18-19,
281, 294-297
emotional intelligence, 11,
254
emotions, 9, 19, 59-60, 71, 77, 133, 136, 153, 240, 246, 252, 254,
283, 295
entrenched culturgen, 100, 102, 242-243
entrenched gene, 39, 43, 78, 241-242
enslavement, see genetic enslavement
eukaryote, 29
eugenics (see also dysgenia),
175-177, 257, 274-276
evolution, pace of, 37-38, 40-42, 49, 84
ESS (evolutionarily
stable strategy), 30, 92
evolutionary psychology (see
also sociobiology), 9, 276
executive function, 71, 83, 85, 136
existentialism, 12, 246-248, 281-283, 290, 313-314
fanaticism (see religion),
10, 114, 116, 121, 126-130, 166, 240
F=ma
(a=F/m), 16, 18-20, 24, 73-74,
108, 155, 157, 280-285, 293-300
Feng Shui, 23
fiction, 142, 153-154
figure/ground (reversal), 81
flag waving (see also
patriotism), 263
free will, 15, 19, 173, 254, 291, 295-299
frontal lobes (see also
brain), 62, 66-67, 71 (plus another 28 pages)
Gage, Phineas, 71
g factor (IQ), 83
gene frequencies, 114-117,
122-124, 126, 159, 172, 241, 269, 301-302
gene interaction effect
(see pleiotropy and polygene)
genetic enslavement, 2-3,
12, 18, 71, 132, 135, 143-144, 147, 152, 154, 259, 261, 266, 313-314
genotype, 50-52,
54, 59, 99, 123, 144, 165-166, 257
GEP, 50-52, 144, 165
glaciers (glacial, interglacial),
9, 36, 40, 42, 47, 54, 89-91, 97, 101-102, 130, 133-134,
143, 234-235, 306
God (gods), 1, 15-16,
20, 22-25, 82, 117-118, 138-139, 148-149, 155-157, 176, 246, 259, 277, 282,
284-289, 295-297, 300
Greek (philosophers,
civilization), 1, 13, 24, 26, 105, 108, 121, 126, 129, 170, 218, 220-223, 243-244, 250, 254, 284
group (selection, subservience
to, collective, see conformism), 10, 47, 92, 94-95, 114-132, 160-164, 303, 307
HBES (Human Behavior
and Evolution Society), 130, 276
Heisenberg Uncertainty
Principle, 17, 96
Holocene, 10, 42,
48-49, 77, 89-90, 95, 97, 102-103, 130, 136, 143-144, 173, 232-234,
241, 307-308
homosexuality, 94-95
hypocrisy, 2, 265-266
IF/THEN, 52-53
human universals, 86,
261-262, 268
individual selection, 10,
114-115, 118, 122-129, 161-164
individualism vs
genes & groups, 1-3, 5-11, 27-29, 31-32, 104, 114-132, 151, 236, 255, 264, 301-303
infanticide, 55
inward-looking (for Truth), 127,
150, 173-175
immigration, 54, 218
immune (response,
system), 36-37, 41, 48, 50-51, 56-57, 165-166, 169-171, 266, 301-302
inclusive fitness, 7, 29,
31-32, 58, 97, 115, 120
innovation rate, 10-11, 20,
33, 106-107, 167, 209-226, 243
interglacial, see glacial
IQ, 11, 57, 82-83, 108, 136, 139, 141, 176-177
lag, genetic, 42
laterality (see also LBS and
RBS), 67-72, 75-80, 136-139, 150-153
left brain (left brained),
60-72, plus ~38 other pages.
LBS, LB-style (left-brain
style), 85, 103-107, 137-143, 146-149, 174-175,
245
levels for selection,
8, 117-120, 123-125
levels of physical explanation,
282, 293-300
liberation, 2, 11, 77, 103,
109, 116, 121-122, 127, 132, 135, 143, 145, 147, 152-154,
174, 230, 252, 254-255, 276, 283
linkage disequilibrium,
92, 268
literacy rate, 78, 178-182
living wisely, 11, 246-252, 254, 270
longevity (fecundity tradeoff),
see also parasitic castration, 259
loyalty (tribal), 3, 95-96, 101, 116, 119, 121, 126, 228
maladapted, see pre-maladapted
Marxism, 3, 118-119,
146-147, 175, 235
materialism (mechanical), 15,
19, 157, 295
Minoan civilization,
105-106, 121, 129, 160, 244
modularity of mentality, 71,
75, 80, 134, 158
monogamy, 55-56, 169
mutation, 5, 29-30,
38-44, 57, 59, 68, 76, 78, 116, 126, 128, 134, 142, 144,
159, 170, 176-178, 240-241, 259, 275
mutational load, 44-45, 176-178
neural network, 68, 72-76, 134-137, 150, 241
New Age, 28, 215
Newtonian physics,
17-18, 22, 167-168
niche, 9, 35, 47, 63,
67, 98, 100-102, 106, 115-116, 135, 146, 148-152,
247
ontogeny (recapitulates
phylogeny), 76, 95
Oriental thought
(see Asian thought)
outlaw genes, 5-8, 11,
27, 46, 59, 108, 139-140, 209-210, 215-231
parasite, 10, 35, 156-160, 162-165, 170, 218, 261
parasitic castration, 218
patriotism, 139, 147-148,
222-223
phenotype, 37-39,
43, 50-52, 79, 84, 90-91, 118, 120, 139, 154, 157-158, 215-216
philosopher, 6-8,
13, 16, 24, 26, 71, 87, 106-107, 116, 121-122, 145, 154, 166-167, 209, 235-237,
243
Philosophes (Enlightenment),
13, 16, 26, 106, 116, 166, 243
pleiotropy, 5, 11, 38, 43, 84, 201-202, 216-217
Pleistocene, 42,
48, 77, 88-89, 91, 128, 131, 165, 194-196, 266
poetry, 104, 137, 237,
243-245
political correctness (PC),
26, 107, 141, 167-168
polygamy, 55
polygenes, 5, 11, 38-39, 78, 80, 82, 201-202, 216-217
population (of world, crash
scenario), 10-11, 106, 161-162, 171-205
postmodernism (deconstructionism,
see also political correctness), 151
pre-adapted (see also pre-maladapted),
9, 33-37, 46-48, 101-102, 136-137, 266-268
predator, 33, 35, 110, 202
pre-maladapted, 9, 34-35
prokaryote, 29
quantum field theory,
quantum mechanics, quantum physics, 13-14, 16-18, 20, 24,
73, 107, 150-152, 239, 252-253, 257-258
random location principle,
anthropic principle, 186-188, 190-191
reductionism, 2,
9, 13-21, 26-28, 51, 60-61, 72, 74, 80, 107, 112, 115, 119,
136, 150, 201, 203, 240-243, 253-260
regression, genetic, 34-35
religion, 1, 13, 15, 23,
26, 85-86, 121, 123-125, 133, 143, 151-153, 203, 229, 238, 247
religious fanaticism, 10,
116
RAS (reticular activating
system) 67, 74, 77, 81-82, 129, 131
right brain (RB,
right cerebral cortex), 10, 23-24, 26, 30, 67-71, 75-79,
82-83, 85-87, 102-107, 128, 130-137, 141-146, 148, 150-151, 153, 166-167,
201, 205, 207, 242
RBS (right-brain
style), 85-86, 132-138, 141-145, 166-167
RDT (result-driven
thinking, or quando conviene), 86-87
reciprocity, 96
rigid universe, 152,
207-208, 237-239
robot (see automaton)
salvation, 206
selfish gene, 7, 9, 31,
115
sexual cannibalism, 31
Shinto cult, 23
siblings, 7, 58
sickle cell anemia, 263
sociobiology (sociobiologist,
see also evolutionary psychology), 4, 7-9, 28, 75, 107, 109-110, 115, 119,
132, 139, 153, 226, 236, 242, 274
spider, Australian redback,
31, 59
spirits, 13, 15, 20, 22-26,
85, 133-138, 150-151, 166, 197, 244, 255, 258, 260
stimulus/response (SR), 51, 158
super-tribes, 10, 96, 111, 116-117,
120, 138, 195
taboo, 92-99,
138, 168, 170, 202-203, 217, 226, 229
takeover males, 54-55, 58, 224
toolmaker, 88-93,
96-97, 137-138, 196
tribe (tribalism), 90-96, 99-103, 194-199, plus 53 others.
two cultures, 10, 85-86,
136-138, 141, 166
utopia, 11, 27, 217, 232-239
virus, 31, 37, 50-51,
162, 261-263
Voltaire, 106, 244
warfare (inter-tribal),
10, 35, 53-54, 89-90, 111, 116, 121, 131, 146, 168, 235
warrior, 89-99,
104-106, 126, 138-139, 160-161, 196-197, 222-224
wealth disparity, 159-160
weapon maker (see toolmaker)
Wernicke’s Area, 67-69,
75, 79, 103, 128, 130
8c25,2131